Övervakning av genetisk mångfald
Genetisk mångfald är den ärftliga biologiska variation som alla arter bär på. Havs- och vattenmyndigheten ansvarar för övervakning av genetisk mångfald hos vattenlevande arter.
De senaste resultaten hittar du under rubriken Resultat från övervakning.
Genetisk mångfald
Vad är genetisk mångfald?
Genetisk mångfald är den ärftliga biologiska variation som alla arter bär på. Den kommer ursprungligen från sällsynta nya mutationer men blandas om inför varje ny generation genom sexuell reproduktion. Mycket genetisk variation gör att en art eller population är mer resilient och bidrar med mer ekosystemfunktioner. Stor genetisk mångfald är särskilt viktig under snabba miljöförändringar.
Genetisk mångfald gör det möjligt för populationer av arter att anpassa sig när miljön förändras. Stor genetisk mångfald – en stor genpool – påverkar således både arternas och hela ekosystemens motståndskraft och funktion positivt. När vi dränerar populationer på deras genetiska mångfald förstör vi deras potential för anpassning, och deras långsiktiga överlevnad kommer att äventyras.
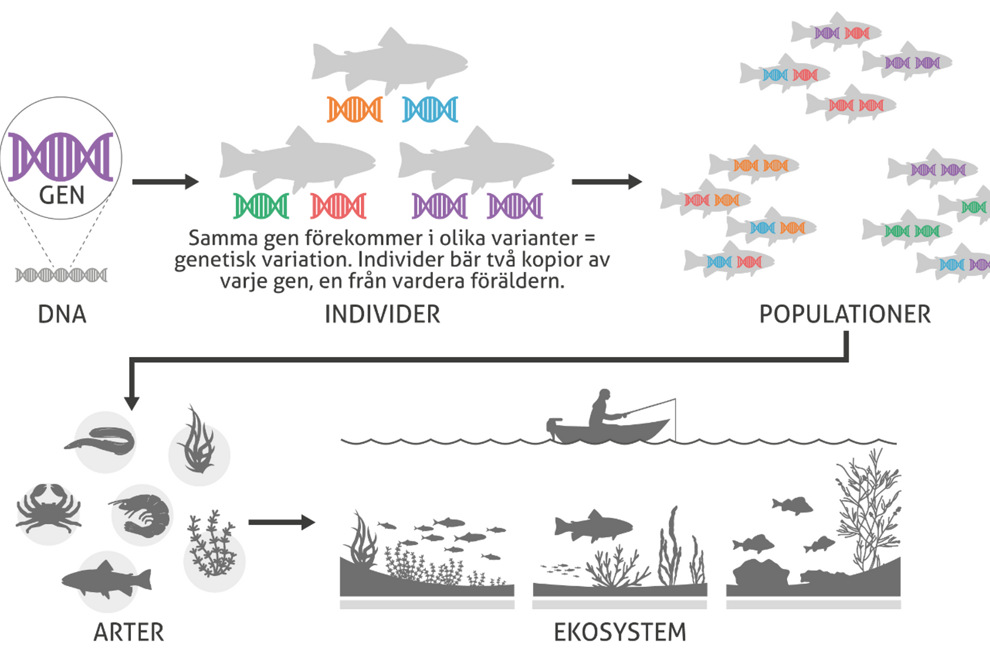
Genetisk mångfald (variation på DNA-nivån) utgör basen av all biologisk mångfald. Färgerna illustrerar olika varianter av en viss specifik gen som orsakas av skillnader i DNA. Ju större färgvariation, desto högre genetisk mångfald i den aktuella genen. Genetisk variation förekommer inom och mellan populationer av samma art. De flesta arter har mellan 10 000 och 20 000 gener i sitt DNA, som alla innehåller ett flertal genvarianter. Illustration: Jerker Lokrantz/Azote.
Varför är genetisk mångfald viktig?
Stor genetisk mångfald är viktig för ekosystemen för att den:
Stor genetisk mångfald hos arter ökar ekosystemens resiliens och funktion, speciellt viktigt är den genetiska variation hos habitatbildande eller reglerande arter i ekosystemet. Förklaringen är att en population med mer genetisk variation har större variation av olika egenskaper. Detta bidrar till fler olika ekosystemfunktioner, såsom ökad produktivitet, ökad motståndskraft mot temporär stress och ökad komplexitet vilket gynnar biodiversiteten av arter i ekosystemet.

Ålgräs är en art som speciellt i Östersjön förökar sig både sexuellt men även genom att bilda kloner. Ålgräsängar som består av många genetiskt olika kloner (hög genetisk mångfald) producerar mer biomassa, utgör livsmiljö för fler andra arter, och är mer motståndskraftiga mot extrema väderförhållanden än ängar med få kloner. Foto: Länsstyrelsen Västra Götaland.
Genetisk mångfald är en förutsättning för lokal anpassning, vilket innebär att populationer av en art kan anpassa sig till olika miljöförhållanden. Finns det exempelvis genetisk variation inom arten med någon genvariant som fungerar bättre i högre temperatur och en annan i lägre temperatur, så har arten en möjlighet att finnas i båda miljöer.
Idag framgår att nästan alla marina arter som undersökts i svenska vatten har lokalt anpassade populationer. Detta trots ofta god förmåga att spridas över stora områden. ch östersjömussla exempel på arter med lokal anpassning runt svenska kusten. I de flesta fall har arternas populationer i Östersjön anpassats till framförallt en lägre salthalt.
Den östra populationen av torsk i Östersjön har exempelvis utvecklat egenskaper som gör att de kan leva och föröka sig i Östersjöns låga salthalter. Bland annat har deras ägg en bättre flytkraft än andra torskars ägg, vilket gör att de inte sjunker i den lägre salthalten. Torsk som inte har denna anpassning kan inte föröka sig i denna miljö. De kan alltså inte på kort tid ersätta östersjötorsken om denna populationen försvinner. Detta leder till en lokal förlust av en art som det kan ta mycket lång tid för naturen att ersätta.
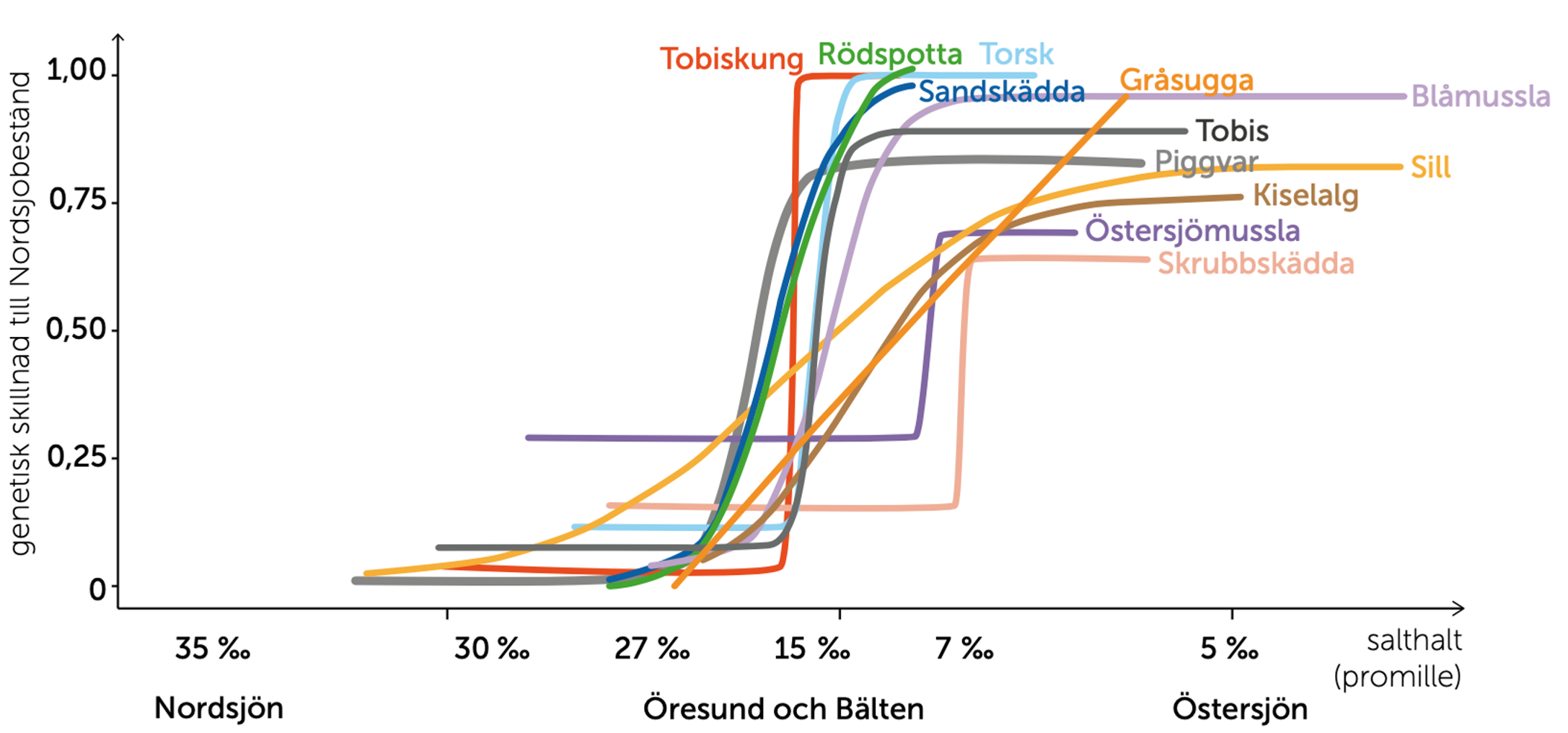
De marina arter som har sin utbredning in i Östersjön är alla mycket vanliga arter i västerhavet och kan redan har burit på stor genetisk variation som möjliggjort en anpassning in i Östersjöns bräckta vatten. Idag uppvisar dessa arter stora genetiska skillnader mellan populationerna i Nordsjön och i Östersjön. Skillnaderna speglar åtminstone delvis en anpassning till Östersjöns lägre salthalt. Källa: Havsutsikt nr 1, 2021
Med större genetisk mångfald (mer genetisk variation) finns en bättre potential för en art eller en population att anpassa sig till pågående och framtida miljöförändringar. Stora populationer har betydligt mer genetisk variation än små populationer. Arter som finns etablerade i flera olika miljöer med lokalt anpassade populationer innehåller betydligt mer genetisk variation än arter som bara finns i en typ av miljö. Arter eller populationer som genomgått dramatiska minskningar i individantal behöver snabbt öka i storlek igen för att inte tappa en större del av sin genetiska variation.
En population kan innehålla "slumrande" genetisk variation som är obrukbar i den livsmiljö den lever i, men som kan bli livsviktig efter en miljöförändring. En annan möjlighet är att en ny mutation tillför genetisk variation som behövs vid en miljöförändring, men chansen att detta ska hända är oftast mycket liten. Det gäller alltså att bevara så mycket av existerande genetisk variation som möjligt i alla arter då man oftast inte på förhand kan avgöra vilka genvarianter som blir viktiga i en ny miljö.
Strandsnäcka (Littorina saxatilis)

Strandsnäckan Littorina saxatilis har anpassat sig till olika miljöer. Blyg krabbsnäcka till vänster och modig vågsnäcka till höger - även personligheterna styrs av gener. Foto: David Carmelet.
På svenska västkusten finns en strandsnäcka (Littorina saxatilis) som har anpassats till två helt olika miljöer, stränder med krabbor och stränder med stark vågexponering.
Men försök i naturen visar att de stora, blyga och tjockskaliga “krabbsnäckorna” kan anpassa sig till vågmiljöer och få samma egenskaper som de små, modiga och tunnskaliga “vågsnäckorna” efter bara 30 år av naturlig selektion. Förklaringen är att även krabbsnäckorna från början innehåller den genetiska variation som behövs för att leva i en vågmiljö, även om våg-generna är ovanliga bland krabbsnäckor som lever i krabbmiljön.
Läs mer i artikeln från Science Advances: Ett 30-årigt fältexperiment på marina sniglar (artikeln är på engelska).
Hur bevaras genetisk mångfald?
Genetisk mångfald försvinner över tiden som en följd av naturligt urval som sorterar bort alla utom den bästa varianten av en gen. Även slumpen orsakar förluster av genetisk variation. Det mest effektiva sättet att motverka förlust av genetisk mångfald är att upprätthålla stora populationer som är sexuellt reproducerande och i kontakt med andra populationer genom utbyte av individer.
Små och isolerade populationer kommer snabbt att förlora genetisk variation på grund av slumphändelser samtidigt som inavelsgraden i populationerna ökar. Detta resulterar i lägre anpassningsförmåga, förlust av motståndskraft och svag potential för långsiktig överlevnad. Populationer som bara förökas asexuellt har mist sin förmåga till snabb anpassning och även förmågan att bli av med skadliga mutationer.
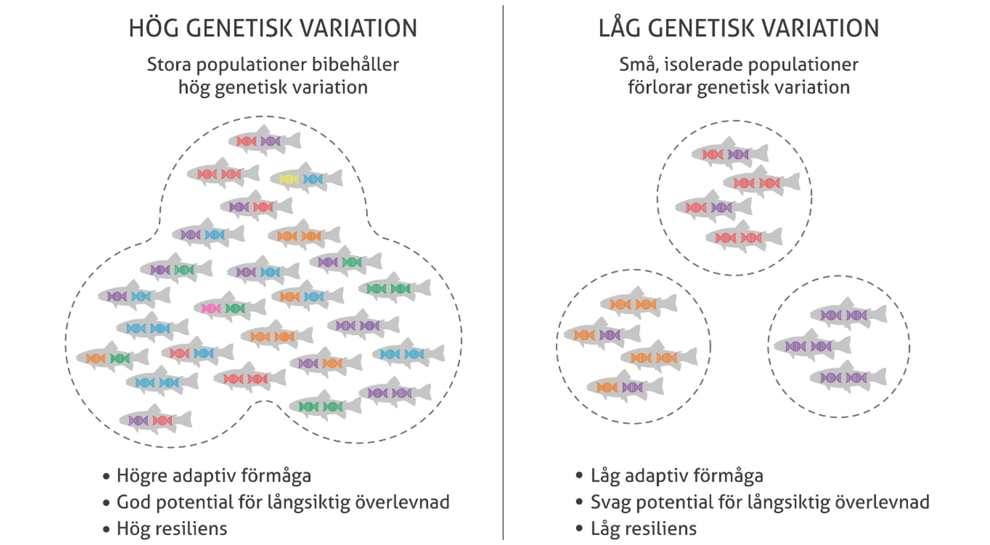
Stora populationer innehåller mer genetisk variation än små genom fler nya mutationer och plats för mer genetisk variation än i små populationer. Illustration: Jerker Lokrantz/Azote.
Vad består genetisk variation av på DNA-nivån?
Den mest basala nivån av biologisk mångfald är den genetiska variationen som finns inom och mellan individer i en population. Denna variation uppkommer som en följd av att olika individer har olika sekvenser av kvävebaser i DNA-molekylerna (kromosomerna). En del av denna variation är det vi kallar gener, vilka påverkar individens egenskaper och funktioner. Utöver detta finns DNA sekvenser som kan vara involverade i DNA-molekylens 3D-struktur eller vara inblandade i genregleringen, det vill säga att “slå på” eller “slå av” gener. Vissa DNA-sekvenser kan också vara historiska rester, som till exempel gener som inte längre fungerar på grund av skadliga mutationer och denna del av DNA saknar direkt funktion. Dessa mer eller mindre "neutrala" DNA sekvenser kan användas för att utröna släktskap, genetiskt utbyte mellan populationer av en art ("konnektivitet"), och storleken av genetiska populationer (s.k. effektiv populationsstorlek).

Hot mot genetisk mångfald
Det finns fyra typer av huvudprocesser som kan leda till förlust av genetisk mångfald:
- Genetiskt särpräglade populationer utrotas
- Genetisk variation inom enskilda populationer förloras på grund av liten storlek, vilket också leder till ökad inavel (liten genetiskt effektiv storlek, Ne)
- Selektion via människans aktiviteter utarmar populationernas genetiska mångfald
- Spridning av genetiskt främmande populationer till områden där naturliga populationer finns och där hybridisering leder till att vilket kan leda till att genetiska anpassningar till lokala ekologiska förhållanden går förlorade.
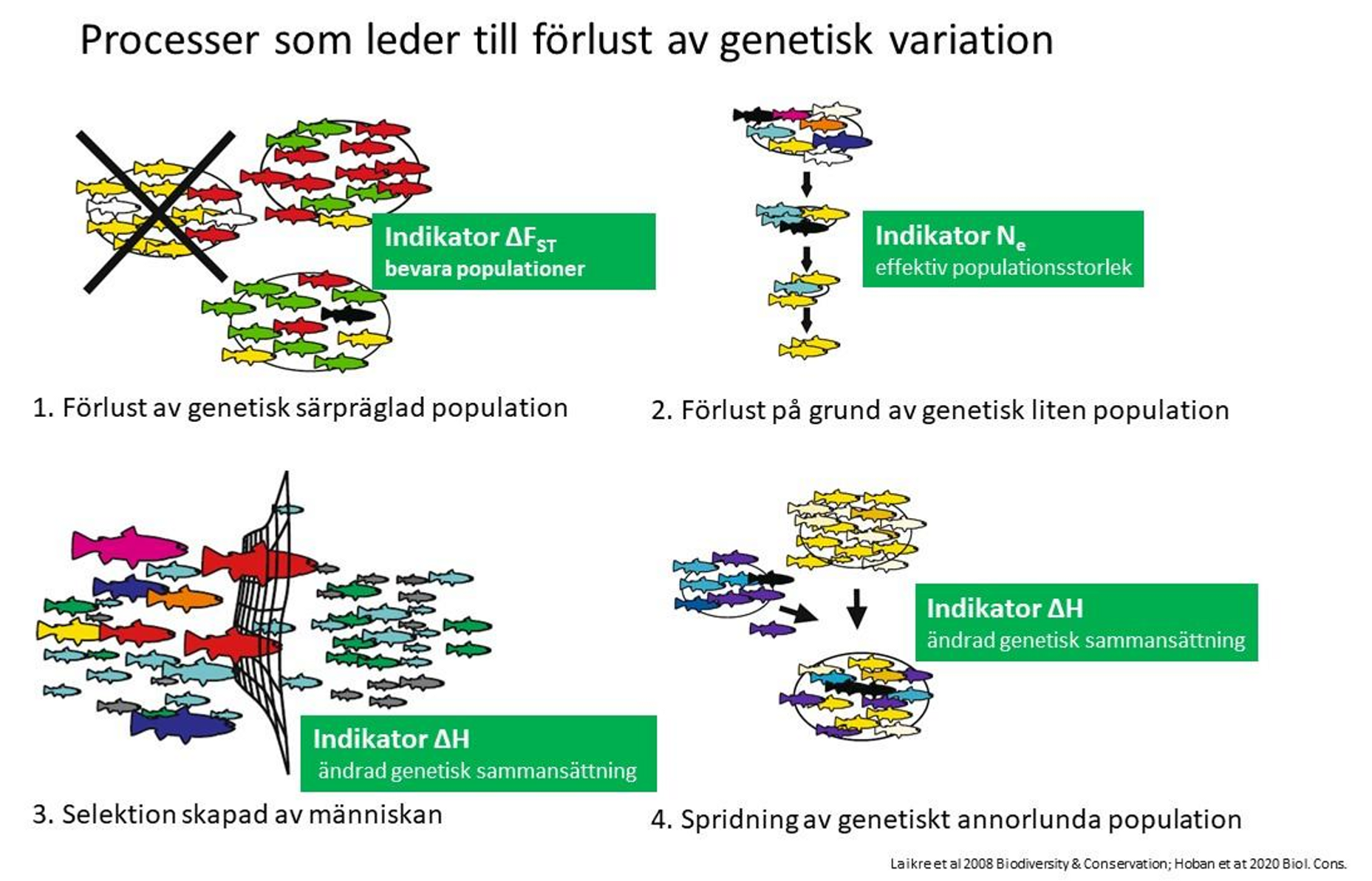
De fyra huvudsakliga hoten mot genetisk mångfald och vilka av dessa processer som de tre indikatorerna som används i övervakningen fokuserar på. Illustration: Laikre et al. 2008 (modifierad)
Läs mer om de tre indikatorerna som används i övervakningen.
Övervakning av genetisk mångfald
Vad är övervakning av genetisk mångfald?
Populationsgenetisk kartläggning
För att övervaka genetisk variation krävs först en grundläggande kartläggning av artens genetiska populationsstruktur. Detta innebär att vävnad från arten samlas in över det geografiska område som ska kartläggas och att DNA-baserade analyser utförs för att undersöka hur den genetiska variationen fördelar sig inom och mellan populationer över området.
En kartläggning är det första steget för att förstå hur mycket genetisk variation som förekommer inom och mellan populationer av en art i ett visst geografiskt område. Till exempel behövs detta för att identifiera och särskilja genetiskt distinkta lokala populationer. Genom att studera hur den genetiska variationen är fördelad kan man också bedöma om ett övervakningsprogram behövs och i så fall hur det ska utformas. För vissa arter finns redan sådan grundläggande kunskap, medan den för andra arter kan vara obefintlig. En första kartering avslöjar också om arten reproducerar sig mer eller mindre asexuellt eller enbart sexuellt.
Övervakning av genetisk mångfald över tid hos populationer och arter
För att så tidigt som möjligt kunna upptäcka hoten mot den genetiska mångfalden behövs övervakning över tid. Detta är särskilt viktigt för arter som utsätts för intensivt fiske eller skörd, används i restaurering eller är föremål för utsättning – insatser som vi vet kan vara kopplade till genetiska förändringar. Övervakning är också nödvändig för arter och populationer som hotas av lokala eller globala miljöförändringar, inklusive effekter av föroreningar eller klimatförändringar. Analyser av genetiska förändringar över tid kan genomföras i de fall arkiverad vävnad eller genetiska data från tidigare studier finns tillgängliga.
I praktiken genomförs genetisk övervakning genom laborativa analyser av särskilt utvalda delar i cellkärnans DNA där det förekommer variation, framförallt med genetiska markörer, så kallad SNPs (Single Nucleotide Polymorphisms) som antingen kan fördelas mer eller mindre slumpmässigt över hela genomet (DNA strängarna) eller lokaliseras till specifika förutbestämda platser med hjälp av en SNP-panel (ibland kallad SNP-chip). Allt vanligare blir också att hela genomet sekvenseras för alla individer som provtas, och i vissa fall kan selektiva genetiska förändringar detekteras. Även med ett relativt begränsat antal SNPar (<1000) eller med s.k. mikrosatelliter (DNA-sekvenser som ofta innehåller mycket genetisk variation), kan man kartlägga genetisk populationsstruktur, följa genetiska förändringar över tid samt skatta effektiv populationsstorlek, genflöde, släktskap,och inavelsgrad. Metodutvecklingen går snabbt och detta motiverar arkivering av vävnadsprover i provbanker där historiska prover kan återanalyseras med än bättre metoder i framtiden.
Inte eDNA
Det är viktigt att inte förväxla dessa populationsgenetiska analysmetoder med eDNA (miljö-DNA), som primärt används för att identifiera vilka arter som förekommer i ett specifikt område och, i vissa fall, även kan ge information om den relativa mängden biomassa hos dessa. I dagsläget kan eDNA inte användas för att studera genetisk mångfald eller genetiska processer inom arter och populationer.
Indikatorer för genetisk mångfald
Indikatorer är mätvärden som indikerar trender i tillståndet i, i detta fall, genetisk mångfald. Eftersom genetisk variation förekommer inom, respektive mellan, populationer av arter behövs indikatorer som mäter båda dessa typer av genetisk variation över tid.
Tre indikatorer som använder sig av genetiska data från kartläggningar av DNA används sedan 2020 i Sverige. De presenterades första gången vetenskapligt av Andersson et al. (2022) och har därefter använts för flera arter.
- Indikator Ne: mäter den genetiskt effektiva storleken hos populationer. Det är en standardparameter inom populationsgenetik som kopplar till en populations förmåga att utveckla nya anpassningar (adaptiv förmåga). Riktvärde för en god adaptiv potential är Ne≥500 och detta är också en huvudindikator inom ramen för FNs konvention om biologisk mångfald. I Sverige används fler tröskelvärden där Ne < 50 leder till röd varning pga mycket låg adaptiv förmåga.
- Indikator ΔH: mäter skillnader i genetisk variation inom population över tid. Flera olika typer av mätvärden används (se figur).
- Indikator ΔFST: fokuserar på genetisk variation mellan populationer. Den följer i vilken utsträckning genetiskt särpräglade populationer (vanligen lokalt anpassade populationer) bibehålls över tid. Den mäter även i vilken utsträckning graden av genetisk skillnad mellan bestånd bibehålls och därmed om det genetiska utbytet (konnektiviteten) förändras över tid.
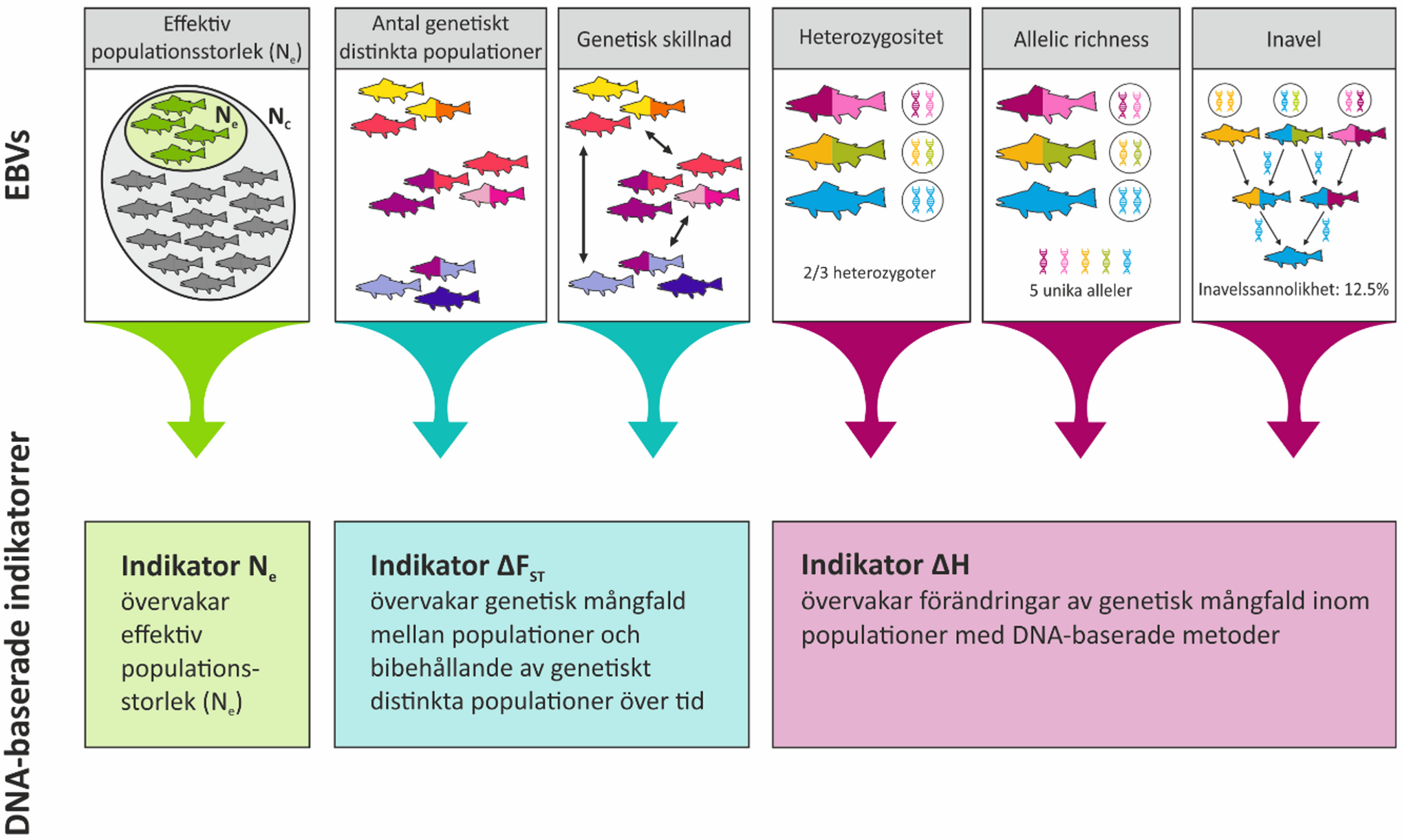
De tre indikatorerna för genetisk mångfald som utvecklats och används i Sverige är alla baserade på centrala och standardmässiga DNA-mått på genetisk variation. Dessa mått kallas även EBVs - Essentiella Biodiversitets-Variabler. Genetisk skillnad mellan bestånd mäts med parameter FST. Inom population mäts genetisk variation bland annat som heterozygositet och allelic richness (ett mått på antalet genvarianter per gen. Inavel kan mätas med vissa DNA-metoder och är en viktig parameter att övervaka. Illustration och copyright: Anastasia Andersson, Stockholms universitet.
Essentiella biodiversitetsvariabler, EBV
I de tre indikatorerna följs trender i olika DNA-baserade mått på genetisk variation. Dessa kallas ibland för essentiella biodiversitetsvariabler för genetisk variation (EBVs; Essential Biodiversity Variables; https://onlinelibrary.wiley.com/doi/10.1111/brv.12852). I figuren exemplifieras flera EBVs som ingår i de DNA-baserade indikatorerna som HaV använder.
För att tolka statistiskt säkerställda förändringar i mätvärden (t.ex. EBVs) som ingår i indikatorerna tillämpas tröskelvärden. Dessa bygger i sin tur på vetenskapligt föreslagna riktlinjer. Gränsvärdena illustreras nedan. Mer information finns i Andersson et al. (2022) samt i manualerna för respektive art (se under respektive art som övervakas genetiskt).
Bildtext: De tre indikatorerna för övervakning av genetisk mångfald (ΔH, Ne, ΔFST) och de gränsvärden som föreslagits och hittills använts i övervakningen. I ΔH-indikatorn ingår flera mått på genetisk variation (EBVs), vilka som används beror bland annat på vilken typ av markör som undersöks (se Andersson et al. 2022; Kurland et al. 2023, Saha & Kurland et al. 2024). HE=förväntad heterozygositet, HO=observerad heterozygositet, AR=allelic richness, NA=antalet alleler, PL=proportionen polymorfa loci, π=nukleotiddiversitet, FROH=inavelsgrad.
Gränsvärden för indikatorer för genetisk mångfald
För att tolka statistiskt säkerställda förändringar i mätvärden (t.ex. EBVs) som ingår i indikatorerna tillämpas tröskelvärden. Dessa bygger i sin tur på vetenskapligt föreslagna riktlinjer. Gränsvärdena illustreras nedan. Mer information finns i Andersson et al. (2022) samt i manualerna för respektive art (se under respektive art som övervakas genetiskt).
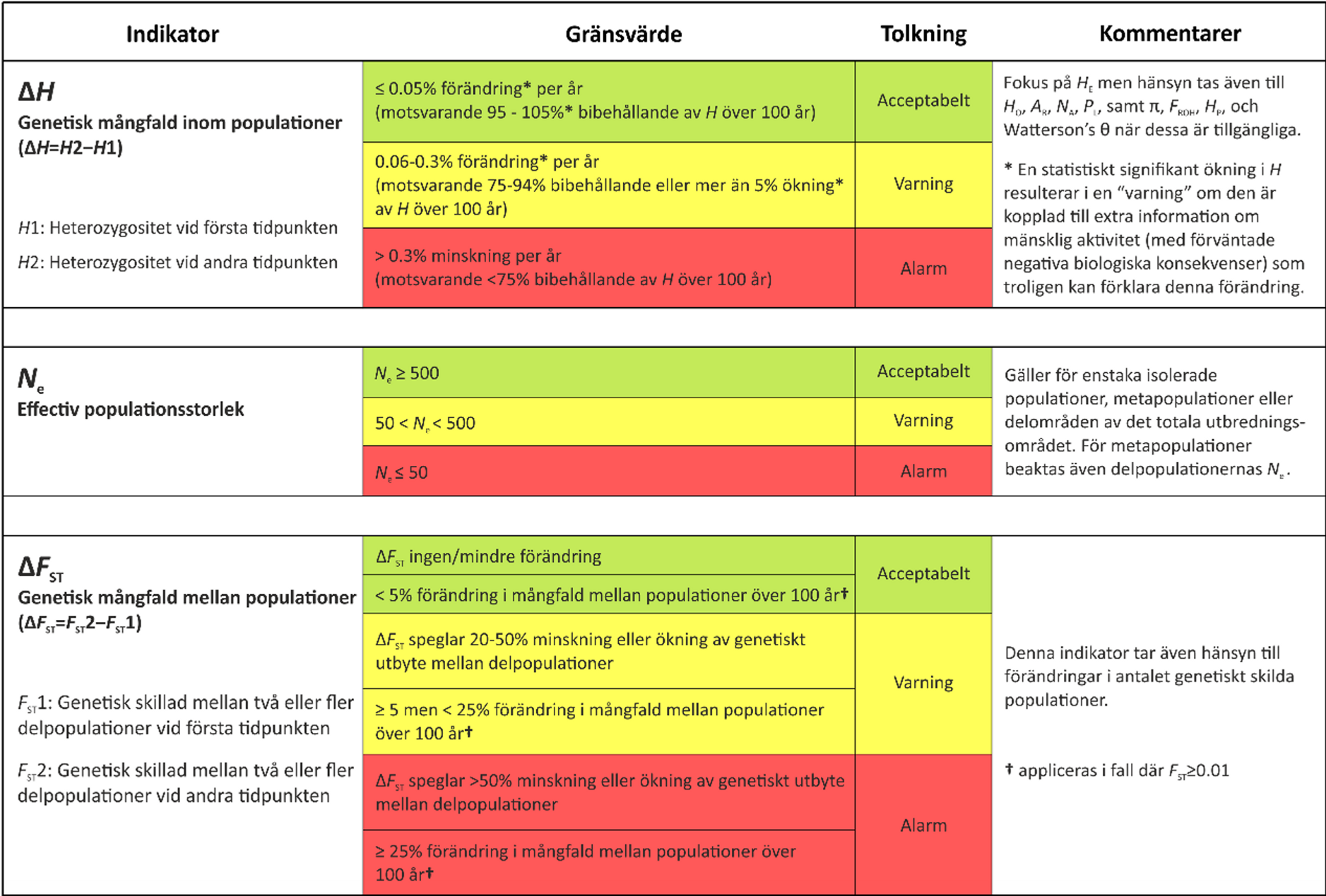
De tre indikatorerna för övervakning av genetisk mångfald (ΔH, Ne, ΔFST) och de gränsvärden som föreslagits och hittills använts i övervakningen.
I ΔH-indikatorn ingår flera mått på genetisk variation (EBVs), vilka som används beror bland annat på vilken typ av markör som undersöks (se Andersson et al. 2022; Kurland et al. 2023, Saha & Kurland et al. 2024). HE=förväntad heterozygositet, HO=observerad heterozygositet, AR=allelic richness, NA=antalet alleler, PL=proportionen polymorfa loci, π=nukleotiddiversitet, FROH=inavelsgrad.
Hur kunskap om genetisk mångfald omsätts i praktisk förvaltning
Fiske
Analyser av genetisk mångfald och struktur är den mest pålitliga, och ibland enda, metoden för att identifiera och särskilja lokala fiskpopulationer.
Att förvalta blandbestånd som om de består en enda genetiskt homogen population innebär särskilt risker för de mindre produktiva populationer som beskattas. Exempel på detta är överfiske i havet av lax från mindre älvar i Östersjön samt utrotningen av små lokala kustbestånd av torsk längs svenska västkusten som idag förvaltas som en del av det stora nordsjöbeståndet.
Populationer som utsätts för omfattande fiskeridödlighet löper stor risk för förlust av genetisk variation och genetiska förändringar som leder till en lägre naturlig tillväxt (långsamväxande individer gynnas) och en sämre produktion av fiskbiomassa.
Innan någon art av fisk, skaldjur eller makroalg skördas kommersiellt, bör en kartläggning av artens genetiska populationsstruktur genomföras. Regelbunden genetisk övervakning kan behövas vid alltmer omfattande fiskeriverksamhet, men är särskilt viktig i samband med blandbeståndsfiske.
Utsättning av fisk och skaldjur
Vid en utsättning släpps individer uppfödda i fångenskap (eller förflyttade) ut i naturen. I Östersjön genomförs exempelvis varje år omfattande utsättningar av odlad lax, öring och sik för att kompensera fisket för vattenkraftsutbyggnadens effekter på älvarna. Odling och utsättning kan dock reducera eller förändra den genetiska mångfalden hos vilda populationer, vilket kan försämra deras anpassningsförmåga till miljöförändringar samt äventyra deras långsiktiga överlevnad.
Utsättningar bör ersättas av en förvaltning som strävar efter att maximera den vilda produktionen. Utsättningar bör i första hand användas i bevarandesyfte (se Restaurering). Det är viktigt att noggrant följa upp genetiska effekter av en utsättning för att säkerställa att inte den genetiska mångfalden hotas.
Arter som bygger upp viktiga livsmiljöer kan skyddas genom olika typer av förvaltning såsom naturreservat och nationalparker, restriktioner för exploatering eller fiske. De prioriteringar som behöver göras bör bygga på kunskap om de olika populationernas genetiska mångfald och graden av konnektivitet. För en del arter kan graden av genetisk variation skilja mellan närbelägna populationer. Detta gäller inte minst arter som växlar mellan sexuell och klonal förökning (till exempel ålgräs, blåstång i Östersjön och ögonkorall).
Innan prioriteringar görs vid skydd av habitatbildande arter bör de olika populationernas genetiska variation samt dess konnektivitet kartläggas. Generellt bör populationer med högre grad av genetisk variation och högre konnektivitet skyddas i första hand, och sexuellt reproducerande populationer bör skyddas framför de som är dominerade av ett fåtal kloner.
Förlust av en lokal population av en mer eller mindre stationär art kan ibland åtgärdas genom restaurering. (Forskning pågår för arter som ålgräs, ögonkorall, torsk och möjligheterna för framgångsrik restaurering kommer sannolikt öka inom en snar framtid.) I de fall utsättning av individer från andra bestånd planeras bör först en mycket noggrann kartläggning genomföras för att givarpopulationen ska vara så representativ som möjligt i förhållande till den population som ska restaureras. Faktorer att ta hänsyn till är framförallt genetisk likhet men även egenskaper i specifika gener, t.ex. gener för stationär eller migrerande livsstil hos torsk.
Restaurering av förlorade lokala populationer eller stöd till genetiskt utarmade populationer bör i första hand ske med individer från närbesläktade populationer och med liknande genetiska egenskaper.
Flera EU-direktiv rör frågor som kopplar till genetisk mångfald och där HaVs övervakningsinformation kan användas i Sveriges nationella rapportering. Detta gäller till exempel Artikel 17 om Art- och habitatdirektivet där information från genetisk övervakning visar på i vilken utsträckning potential för långsiktig överlevnad och anpassningsförmåga föreligger. Design, skötsel, uppföljning och rapportering av Natura 2000-områden kan förbättras genom att inkludera genetisk information som kan belysa i vilken utsträckning specifika områden bidrar till att säkerställa genetisk konnektivitet inom arten.
Havsmiljödirektivet fokuserar på en god miljöstatus i havsmiljö och där är genetisk mångfald en nyckelfaktor för populationers och arters ekosystemfunktion och överlevnadsförmåga liksom för genetiskt utbyte mellan skyddade områden. Vattendirektivet fokuserar bland annat på miljökvalitetsnormer i vattenmiljö vilket inkluderar god konnektivitet. Övervakning av genetisk variation ger en god bild av biologisk konnektivitet i vattenmiljöer och kan därför användas i rapportering och i förvaltning.
Implementeringen av EU:s nya förordning om restaurering av natur kommer att stöttas av kunskap och övervakning av genetisk mångfald inklusive användande av de föreslagna indikatorerna. Främst gäller detta uppföljning och utvärdering av insatser för att återskapa konnektivitet. Implementering av ett flertal artiklar i förordningen kommer att stödjas av genetisk information från HaVs övervakningsinsatser.
Resultat från övervakning av genetisk mångfald kan användas i implementering och rapportering av EUs direktiv och regelverk som kopplar till biodiversitet och biologisk konnektivitet.
Genetisk mångfald är en viktig aspekt i arbetet med att ompröva vattenkraftens miljövillkor inom den nationella planen (NAP). Vattenkraftsutbyggnad påverkar akvatiska arters förekomst, produktion och spridning genom förändrade habitat, ändrade flödesförhållanden och minskade möjligheter till vandring uppströms och nedströms. Ur ett genetiskt perspektiv förväntas minskade och mer isolerade lokala populationer (fragmentisering) leda till ökad inavel och förlust av genetisk variation. Borttagande av vandringshinder och konstruktioner som möjliggör passage förbi hinder (t.ex. omlöp) kan bör därför förväntas komma att öka den genetiska variationen och minska graden av inavel. På detta sätt fås en positiv effekt på tidigare isolerade bestånd av fisk, musslor och andra sötvattensorganismer.
Vi har idag dålig kunskap om vilka negativa genetiska effekter som drabbar akvatiska arter när vattendrag regleras. Grundläggande genetiska kartläggningar behöver genomföras för arter som flodpärlmussla och vissa fiskar.
FN:s konvention om biologisk mångfald (Convention on Biological Diversity, CBD; www.cbd.int) antog 2022 en ny global biodiversitetsstrategi som sätter starkare och tydligare mål för genetisk mångfald än tidigare.
Tillståndsmålet till 2050 är att populationer (bestånd) av arter ska vara tillräckligt stora för att säkerställa att genetisk variation bibehålls på en nivå som säkerställer populationens adaptiva potential. Åtgärdsmålet till 2030 är att insatser ska vidtas som säkerställer att genetisk variation både inom och mellan populationer bibehålls och att insatser görs för att förbättra läget så att anpassningsförmågan hos bestånden (populationerna inom arter) når hög adaptiv förmåga.
För att övervaka att målen nås ska en huvudindikator som mäter andelen populationer inom arter som når Ne>500 användas. En kompletterande indikator som följer andelen populationer inom arter som bibehålls över tid kan också användas.
De DNA-baserade indikatorer som HaV använder i sin övervakning ger information till båda dessa globala indikatorer och kan användas i Sveriges rapportering till CBD.
Indikatorvärden för genetisk variation som tas fram för populationer av arter inom HaV:s övervakning kan användas i rapporteringen till Konventionen om biologisk mångfald.
Vikten av att säkerställa genetisk mångfald framgår av flera av den svenska miljömålen. I sju av de 16 miljömålen (Levande sjöar och vattendrag, Hav i balans samt levande kust och skärgård, Myllrande våtmarker, Levande skogar, Ett rikt odlingslandskap, Storslagen fjällmiljö och Ett rikt växt- och djurliv) anges att genetisk variation inom och mellan populationer av naturligt förekommande arter i olika naturtyper ska bevaras på tillräcklig nivå.
Indikatorvärde för genetisk variation inom HaVs övervakning och annan information om genetisk mångfald hos övervakade arter kan användas i nationell rapportering av de nationella miljömålen.
Resultat från övervakning
Detta övervakas idag
Havs- och vattenmyndigheten, HaV, har inlett genetisk övervakning av sju akvatiska arter i samarbete med forskare vid Göteborgs universitet, Stockholms universitet och Sveriges lantbruksuniversitet.
Fem arter av fisk (lax, öring, torsk, sill och röding) ingår i övervakningen därför att de riskerar att påverkas genetiskt av fiske eller andra antropogena aktiviteter, samt har viktiga ekologiska roller.
Övervakningen sker från källa till hav, det vill säga fisk i områden högst beläget i fjällen (öring och röding), via vattendrag och älvar (öring, lax) ut till havet (lax, öring, sill, torsk).
Två habitatbildande arter med stor ekologisk betydelse (ålgräs och blåstång) ingår också.
Andra arter som är prioriterade kandidater för framtida genetisk övervakning är abborre, blåmussla, flodpärlmussla, skarpsill, tånglake och vikaresäl, som är bedömt viktiga ur ett ekosystemperspektiv och/eller kan påverkas genetiskt av förändrade miljöförhållanden.
Under respektive artnamn finns detaljerade resultat från övervakningen.

Ålgräs förekommer ofta som ängar som består av ett varierande antal genetiskt skilda individer. I västerhavet är ängarna oftast rika på olika kloner medan Östersjöns ålgräsängar består av en eller ett fåtal kloner. Foto: Länsstyrelsen Västra Götaland.
Resultat från tidigare genetisk kartläggning och nuvarande övervakningsprogram
Ålgräsets genetiska variation har kartlagts både globalt och runt svenska kusten. Arten ålgräs har sitt ursprung i Stilla Havet och har, som många andra marina arter i norra Atlanten, vandrat in norr om Kanada under perioder av mildare klimat. Som en följd av populationsflaskhalsar i samband med invandring till nya områden har ålgräset i Atlanten betydligt mindre genetisk variation än i Stilla havet.
Tidiga analyser på västkusten gjorda med mikrosatelliter visar att ålgräset är uppdelat i olika lokala bestånd. Inom övervakningsprogrammet för genetisk mångfald har en fördjupad studie genomförts med mer kraftfulla genetiska metoder (DNA sekvensering). Denna studie innefattar även lokaler i Östersjön. Studien visar att västkusten innehåller väsentligt mycket mer genetisk variation än Östersjön. Östersjöns lokaler domineras också av vegetativ förökning.
Analyser av ålgräsprover från hela artens svenska utbredning visar att västkusten har 9 olika bestånd ("cluster"), medan alla lokaler unom Nynäshamn i Östersjön tillhör samma genetiska grupp ("cluster"). Västkustens lokaler karatäriseras också av att de alla innehåller många unika genotyper (kloner), medan i Östersjön har många av lokalerna endast ett fåtal kloner. Bestånden vid Björkö, Krampö, Hornsudde, Bläsinge och Björkenäs består av bara en klon. Illustration: Stefanie Ries
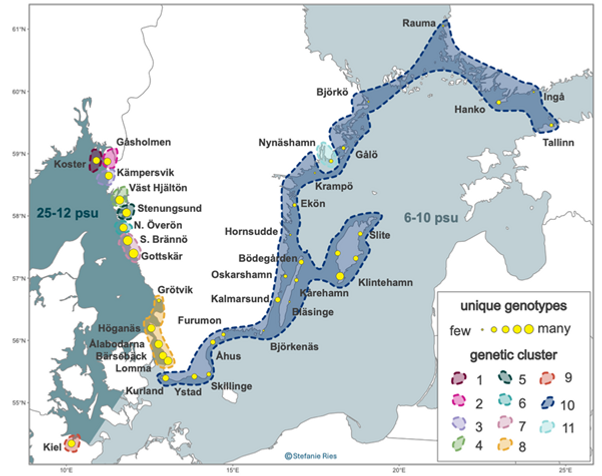
Analyser av ålgräsprover från hela artens svenska utbredning visar att västkusten har 9 olika bestånd ("cluster"), medan alla lokaler unom Nynäshamn i Östersjön tillhör samma genetiska grupp ("cluster"). Västkustens lokaler karatäriseras också av att de alla innehåller många unika genotyper (kloner), medan i Östersjön har många av lokalerna endast ett fåtal kloner. Bestånden vid Björkö, Krampö, Hornsudde, Bläsinge och Björkenäs består av bara en klon. Illustration: Stefanie Ries.
Frågor av särskild relevans
- På västkusten behövs en än mer detaljerad upplösning då alla de nuvarande bestånden har beskrivits från enbart en lokal vardera. Det är viktigt att över tid följa vad som händer med olika lokala bestånd. Inte minst gäller detta i områden där ålgräsets utbredning har reducerats kraftigt under senare tid, som till exempel norr om Nordre Älvs utlopp.
- I Östersjön bör en övervakning ske som framförallt fokuseras på de områden med flera kloner i samma bestånd, för att se om klonrikedomen reduceras över tid.
- Nynäshamnsområdet bör undersökas närmare för att fastställa utbredningen av det bestånd som framstår som en utgrupp i förhållande till Östersjö-klustret.
- Ålgräs restaureras i flera områden på Bohuskusten, och det är viktigt att bevaka att givarbestånden kommer från närområdet.
- Det kan finnas kloner av ålgräs i Östersjön som har bättre tålighet mot uppvärmning än andra kloner.
Vill du läsa mer om arten?
Läs mer på sidan Ålgräsängar.
Vetenskapliga nyckelreferenser
- Ries et al. 2023. Genetic structure and diversity of the seagrass Zostera marina along a steep environmental gradient, with implications for genetic monitoring. Frontiers in Climate 5:1303337.
Den genetiska variationen hos ålgräs undersökt med DNA sekvensering och på prover från hela det svenska utbredningsområdet. Studien visar mycket större genetisk variation på västkusten medan Östersjöbestånden är genetiskt begränsade pga avsaknaden av sexuell förökning. - Reusch et al 2005. Ecosystem recovery after climatic extremes enhanced by genotypic diversity. PNAS 102:2826-2831.
Studien visar att ålgräsängar som bestod av sex kloner återhämtade sig betydligt bättre än de som bara bestod av en klon, efter en period av extrem värmebölja. Studien visar att genetisk mångfald är viktig för ekosystemfunktioner och resiliens. - Duffy et al. Pleistocene legacy structures variation in modern seagrass ecosystems. PNAS 119:e2121425119
Genetisk data från både Stilla Havet och Atlanten att den genetiska variation är mindre i Atlanten än i Stilla Havet och att detta resulterar i sämre tillväxt i Atlanten och mindre variation i egenskaper. Ålgräsets historia med en invandring till Atlanten under vilken genetisk variation gick förlorad påverkar alltså dagens egenskaper hos arten. - Jahnke et al. 2020. Integrating genetics, biophysical, and demographic insights identifies critical sites for seagrass conservation. Ecol. Appl. 30:e02121
Denna studie visar hur olika typer av data kan användas för att pricka in områden som kan prioriteras inom förvaltningen.

Lax. Illustration: Wilhelm von Wright.
Tidigare genetisk kartläggning
De första genetiska studierna av laxens populationsstruktur genomfördes redan på 1980-talet, och idag är laxen en av de mest välstuderade fiskarterna ur genetiskt perspektiv. Inledningsvis användes enzymelektroforetiska markörer (allozymer), följt av studier av variation i mitokondriens DNA (mtDNA). Senare kartläggningar har främst varit baserade på nukleärt nedärvda mikrosatelliter och SNPs.
Laxens förmåga att nästan alltid återvända till sin födelseplats för att leka har resulterat i tydliga genetiska skillnader mellan populationer i olika vattendrag, där graden av differentiering tenderar att öka med det geografiska avståndet. I större älvar kan flera lokala populationer även finnas inom olika delar av samma vattensystem.
I Sverige kan laxen delas in i tre till fyra genetiska populationsgrupper: Vänern, västkusten och Östersjön, där det inom den sistnämnda även finns skillnader mellan populationer i Bottniska viken och Egentliga Östersjön. Dessa mer storskaliga genetiska skillnader återspeglar sannolikt att norra Europa, efter senaste istiden, återkoloniserades av genetiskt distinkta laxar från olika istidsrefugier.
Det faktum att laxen är genetiskt välstuderad, och att vävnadsprover (torkade fjäll, frusen och spritad vävnad) från flera vattendrag finns arkiverade, gör det möjligt att effektivt övervaka genetiska förändringar över olika tidshorisonter och geografiska skalor. De genetiska hot som föreligger mot arten gör denna typ av miljöövervakning särskilt viktig.
Bland de svenska populationerna av östersjölax har genetisk homogenisering kopplad till utsättningsverksamhet observerats över en hundraårsperiod. I Norge har odling av lax för livsmedelsproduktion haft negativa effekter på den vilda laxen, och förrymd norsk odlad lax har även påverkat vilda populationer längs svenska västkusten. I Vänern har de få kvarvarande vilda laxpopulationerna också påverkats av odlings- och utsättningsverksamhet.
Resultat från nuvarande övervakningsprogram
Inom övervakningingsprogrammet för genetisk mångfald har kompletterande genetiska analyser gjorts av arkiverad vävnad från lax från svenska västkusten och Vänern. Dessutom har befintliga data för östersjölax från tidigare forskningsprojekt analyserats statistiskt och utvärderats med avseende på de gränsvärden och kriterier som nyligen tagits fram. Totalt omfattar övervakningen de 17 laxvattendrag/populationer som visas på kartan nedan, varifrån två till sex temporala stickprov har DNA-analyserats.
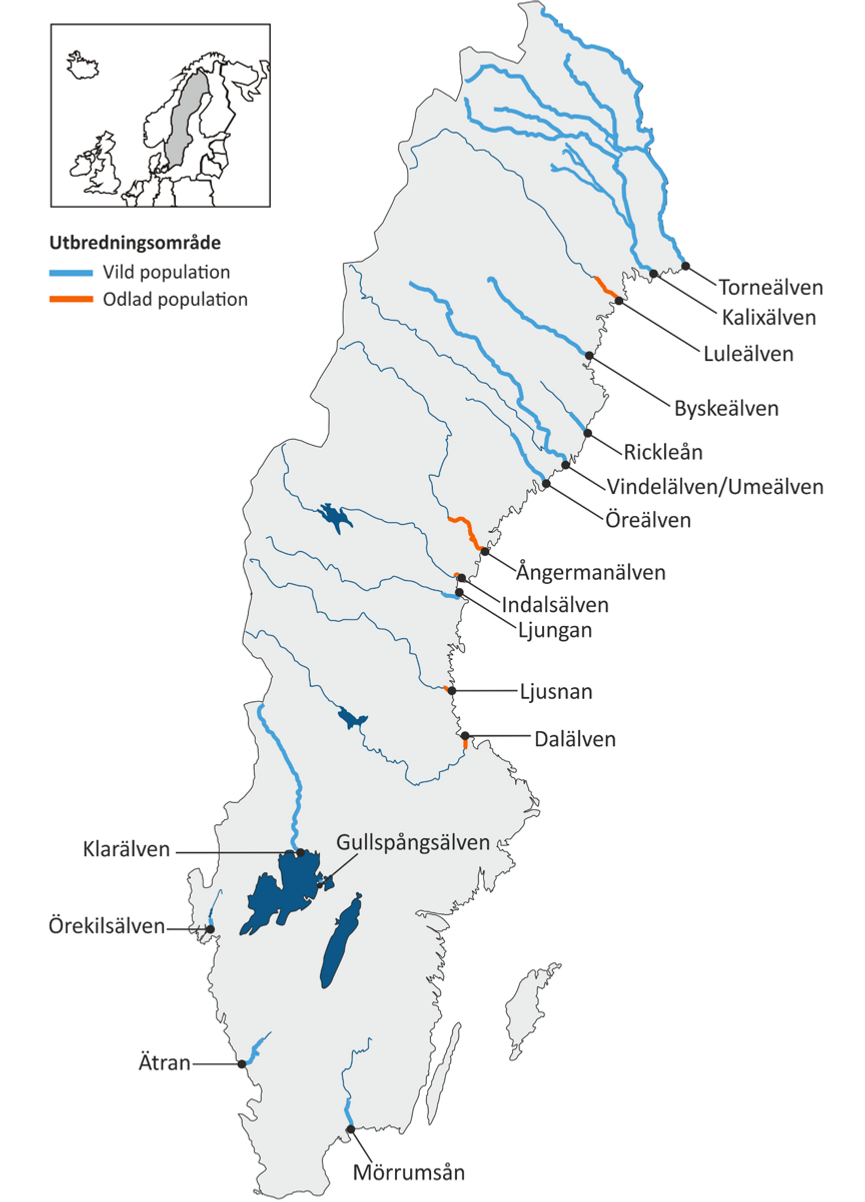
Vattendrag som hittills ingått i den genetiska övervakningen för lax. Tjockare linjer markerar nutida utbredningsområden för idag vilda (blå) respektive kompensationsodlade (röda) populationer. Från Kalixälven, Luleälven, Indalsälven, Dalälven, Mörrumsån och Klarälven finns fler än två temporala stickprov som DNA-analyserats. Illustration: Anastasia Andersson för Andersson et al. in prep.
Resultat och utfall för de genetiska indikatorerna baserade på temporala jämförelser mellan två stickprov från de 17 laxvattendragen är summerade nedan.

Indikator-utslag avseende genetisk variation för laxbestånd från Östersjön (lila), västkusten (blått) och Vänern (orange) med avseende på indikatorerna ΔH, Ne och ΔFST, baserat på jämförelser mellan två temporala stickprov (äldsta och yngsta för populationer med fler än två analyserade prov). De färgade cirklarna visar genetisk status; grön=god, gul=varning och röd=alarm. Pilarna anger den detekterade förändringens riktning, där horisontell pil innebär att ingen förändring har skett; svarta pilar visar på att förändringen är statistiskt säkerställd (p<0.05). Detaljer om använda gränsvärden finns här. Källa: Andersson et al in prep, Johannesson & Laikre 2023).
Varningar har identifierats för flera älvar, främst avseende indikatorn effektiv populationsstorlek (Ne). Endast fyra laxpopulationer (alla i Östersjön) uppvisar Ne >500 (god status, grön). Bland de övriga 13 populationerna har 12 Ne mellan 50 och 500 (varning, gul), medan Gullspångsälven (Vänern) har Ne<50 (alarm, röd).
Fem populationer – Mörrumsån, Ätran, Örekilsälven, Klarälven och Gullspångsälven – får också varning för indikatorn ΔH, tack vare en ökad genetisk variationsgrad, vilket indikerar störningar (ökat genflöde) relaterade till odlingsverksamhet. I linje med detta uppvisar de tre populationsgrupperna (Östersjön, västkusten, Vänern) ökad genetisk homogenisering (indikatorn ΔFST) som överskrider gränsvärdena. Detta medför varning (gul) för samtliga östersjölaxpopulationer och alarm (röd) för de åtta vilda populationerna från samma område. För laxen på västkusten och i Vänern erhåller hela systemet alarm (röd). Under 100 år förväntas de genetiska populationsskillnaderna att minska med 24–29 % för östersjölax, beroende på om samtliga eller endast dagens vilda populationer beaktas. För laxen på västkusten och i Vänern förväntas motsvarande minskningar bli 46 % respektive 40 % (båda alarm, röd) vid en oförändrad förändringstakt.
För de sex populationerna med fler än två temporala stickprov har genetiska förändringar och skillnader i genetisk populationsstruktur mellan olika tidsperioder kunnat studeras närmare, vilket är exemplifierat i följande figur för populationen i Mörrumsån.
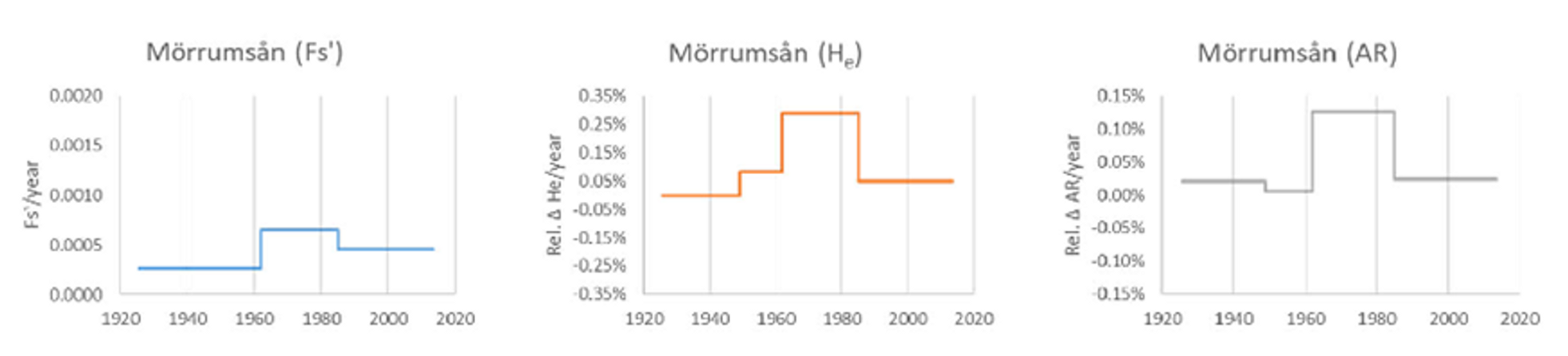
Uppskattad genetisk förändringstakt (per år) under olika tidsperioder i Mörrumsån (fem temporala stickprov: lax provtagen under 1920-, 1940-, 1960-, 1980- respektive 2010-talet). I vänstra delfiguren mäts allelfrekvensförändring (Fs’), där ett högt värde motsvarar en låg effektiv populationsstorlek och vice versa. I den mellersta och högra delfiguren visas förändringar i förväntad heterozygositet (HE) respektive allelrikedom (AR) - två vanliga mått på genetisk variationsgrad, där såväl positiva (ökning) som negativa (minskning) värden kan förekomma. Av samtliga figurer framgår att den genetiska förändringstakten var störst mellan 1960- och 1980-talet.
De största genetiska förändringarna verkar ha inträffat mellan 1960- och 1980-talet, medan förändringstakten har varit lägre både före och efter denna period. Den största påverkan har överlag observerats hos kompensationsodlade bestånd samt inom populationerna i Mörrumsån (Östersjön) och Klarälven (Vänern), medan den vilda laxen i den större Kalixälven (Östersjön) uppvisar betydligt mindre förändringar.
Resultaten indikerar att oönskade genetiska förändringar huvudsakligen ägt rum under tidigare decennier, då kompensationsodlingen etablerades, laxar flyttades mellan odlingar och stödutsättningar genomfördes. Tidigare låga effektiva populationsstorlekar i det vilda kan dessutom ha förstärkt effekterna av genetiskt avvikande individer. För laxen på västkusten saknas ännu lika detaljerade resultat, men den nyligen konstaterade förekomsten och påverkan av förrymd norsk odlad lax på vilda populationer tyder på en fortsatt genetisk påverkan.
Frågor av särskild relevans
Ovanstående varningssignaler indikerar att utsättningsverksamhet har påverkat flera av laxbestånden. Resultaten, vilka kompletterar och stärker slutsatser från tidigare studier, visar att de enskilda bestånden är genetiskt sårbara och att i genomsnitt 20%, 37% respektive 25% av den genetiska variationen mellan bestånden längs östersjökusten, västkusten respektive i Vänern har gått förlorad.
Även om det finns indikationer på att mycket av dessa oönskade genetiska förändringar kan ha ägt rum under tidigare årtionden, när situationen för flera av de vilda laxbestånden var sämre än idag och medvetenheten lägre om att man bör värna artens genetiska mångfald, behöver dessa signaler beaktas inom dagens laxförvaltning.
Vidare bör den genetiska homogenisering som observeras och tar sig uttryck i förändrad genetisk variation inom population och förlust av genetisk särprägel hos bestånden (minskad mellankomponent av genetisk variation) ånyo föranleda överväganden kring de storskaliga utsättningar för lax som fortgår, trots att genetiska problem förknippade med verksamheten har påpekats av forskare sedan 1980-talet.
Vill du läsa mer om arten?
Läs mer på sidan Lax.
Vetenskapliga nyckelreferenser
Laxens genetiska populationsstruktur
Studier av laxens genetiska populationsstruktur på olika geografiska nivåer och med olika molekylära metoder och markörsystem (i kronologisk ordning):
- Ståhl G. 1981. Genetic differentiation among natural populations of Atlantic salmon (Salmo salar) in Northern Sweden. Ecological Bulletins 34:95-105.
- Ståhl G. 1983. Differences in the Amount and distribution of genetic variation between natural populations and hatchery stocks of Atlantic salmon. Aquaculture 33:23-32.
- Ståhl G. 1987. Genetic population structure of Atlantic salmon. In: Ryman N, Utter F (eds) Population Genetics & Fishery Management. University of Washington Press, Seattle and London.
- Koljonen M-L, Jansson H, Paaver T, Vasin O, Koskiniemi J. 1999. Phylogeographic lineages and differentiation pattern of Atlantic salmon (Salmo salar) in the Baltic Sea with management implications. Can. J. Fish. Aquat. Sci. 56: 1766–1780.
- Nilsson J, Gross R, Asplund T, Dove O, Jansson H, Kelloniemi J, Kohlmann K, Löytynoja A, Nielsen EE, Paaver T, Primmer CR, Titov S, Vasemägi A, Veselov A, Öst T, Lumme J. 2001. Matrilinear phylogeography of Atlantic salmon (Salmo salar L.) in Europe and postglacial colonization of the Baltic Sea area. Molecular Ecology 10, 89-102.
- Säisä M, Koljonen M-L, Gross R, Nilsson J, Tähtinen J, Koskiniemi J, Vasemägi A. 2005. Population genetic structure and postglacial colonization of Atlantic salmon (Salmo salar) in the Baltic Sea area based on microsatellite DNA variation. Canadian Journal of Fisheries and Aquatic Sciences 62:1887-1904.
- Bourret V, Kent MP, Primmer CR, Vasemägi A, Karlsson S, Hindar K, McGinnity P, Verspoor E, Bernatchez L, Lien S. 2013. SNP-array reveals genome-wide patterns of geographical and potential adaptive divergence across the natural range of Atlantic salmon (Salmo salar). Molecular Ecology 22: 532–551.
- Miettinen A, Palm S, Dannewitz J, Lind E, Primmer CR, Romakkaniemi A, Östergren J, Pritchard VL. 2021. A large wild salmon stock shows genetic and life history differentiation within, but not between, rivers. Conservation Genetics 22:35-51. DOI: 10.1007/s10592-020-01317-y
- Miettinen A, Romakkaniemi A, Dannewitz J, Pakarinen T, Palm S, Persson L, Östergren J, Primmer CR, Pritchard VL. 2024. Temporal allele frequency changes in large-effect loci reveal potential fishing impacts on salmon life-history diversity. Evolutionary Applications. 17:e13690.
https://doi.org/10.1111/eva.13690
Fokus på laxens genspridning
Artiklar och rapporter med fokus på genspridning från odlad, utsatt lax och andra hot mot laxens genetiska mångfald (i kronologisk ordning):
- Ståhl G, Ryman N. 1987. Lax och öring i Vänernområdet. En populationsgenetisk analys. Naturvårdsverket Rapport 3357.
- Vasemägi A, Gross R, Paaver T, Koljonen ML, Nilsson J. 2005. Extensive immigration from compensatory hatchery releases into wild Atlantic salmon population in the Baltic Sea: spatiotemporal analysis over 18 years. Heredity 95: 76-83.
- Palm S, Dannewitz J, Johansson D, Laursen F, Norrgård J, Prestegaard T, Sandström A. 2012. Populationsgenetisk kartläggning av Vänerlax. Aqua reports 2012:4. 64 pp.
- Palmé A, Wennerström L, Guban P, Ryman N, Laikre L. 2012. Compromising Baltic salmon genetic diversity conservation genetic risks associated with compensatory releases of salmon in the Baltic Sea. Havs- och vattenmyndighetens rapport 2012:18. ISBN 978-91-87025-19-8
- Palm S, Dannewitz J, Prestegaard T, Östergren J (2013). Laxing och felvandrad lax i Mörrumsån: en genetisk analys. Aqua reports 2013:20. 44 pp.
- Karlsson S, Diserud OH, Fiske P, Hindar K. 2016. Widespread genetic introgression of escaped farmed Atlantic salmon in wild salmon populations. ICES J Mar Sci 73: 2488−2498
- Palm S, Karlsson S, Diserud OH. 2021. Genetic evidence of farmed straying and introgression in Swedish wild salmon populations. Aquaculture Environment Interactions 13:505-513.
DOI: https://doi.org/10.3354/aei00423 - Östergren J, Palm S, Gilbey J, Spong G, Dannewitz J, Königsson H, Persson J, Vasemägi A. 2021. A century of genetic homogenization in Baltic salmon – evidence from achival DNA. Proc. R. Soc. B 288:2023147.
https://doi.org/10.1098/rspb.2020.3147

Öring i akvarium. Foto: Natalie Greppi / Havs- och vattenmyndigheten.
Tidigare genetisk kartläggning
Öringen är en av de mest välstuderade arterna ur genetiskt perspektiv. De första studierna av öringens genetiska populationsstruktur genomfördes redan på 1970-talet, och arten har sedan dess varit fokus för omfattande populationsgenetiska studier. Inledningsvis (1970-talet och framåt) användes proteinelektrofores-teknik (enzymkodande gener studeras via variation i enzymernas laddning, så kallade allozymer), följt av studier av variation i mitokondriens DNA (mtDNA). Därefter har kartläggningar ofta baserats på nukleärt nedärvda mikrosatelliter och SNPs, och på senare år har helgenomsekvensering använts.
Samtliga tekniker ger vid hand att öringen har en mycket stark tendens att bilda genetiskt särpräglade populationer. Även över mycket begränsade geografiska områden bildar öringen genetiskt åtskilda populationer med begränsat, men livsviktigt genetiskt utbyte sinsemellan. Genetiskt åtskilda bestånd som samlever i samma sjö upptäcktes redan på 1970-talet (Allendorf et al 1976; Ryman et al. 1979) och att skillnaden mellan sådana bestånd kan vara mycket stor, med olika nivåer av genetisk variation och inavelgrad har på senare år visats med genomikstudier av samma bestånd som studerades på 1970-1980-talen med allozymer (Saha et al. 2022).
Det faktum att öringen är genetiskt välstuderad, och att äldre vävnadsprover från de tidiga analyserna från 1970-talet finns sparade har gjort det möjligt att effektivt övervaka genetiska förändringar över de senaste decennierna. Längre tidsperioder kan övervakas med hjälp av fjällprover som finns för vissa vattendrag. Vidare driver Stockholms universitet sedan slutet av 1970-talet ett övervakningsprojekt av öring i Hotagens naturreservat. Detta projekt har givit värdefull information om öringens populationsgenetik där kunskapen kunnat ligga till grund för att utveckla indikatorer för övervakning av genetisk mångfald.
Resultat från nuvarande övervakningsprogram
Inom ramen för övervakningsprogrammet av genetisk mångfald valdes initial fokus på flera skyddade områden i Jämtlands län där det dels finns prover från 1970-1980-tal, dels pågår provfisken alternativt forskningsinsatser som möjliggjorde effektiv insamling av nutida prover.
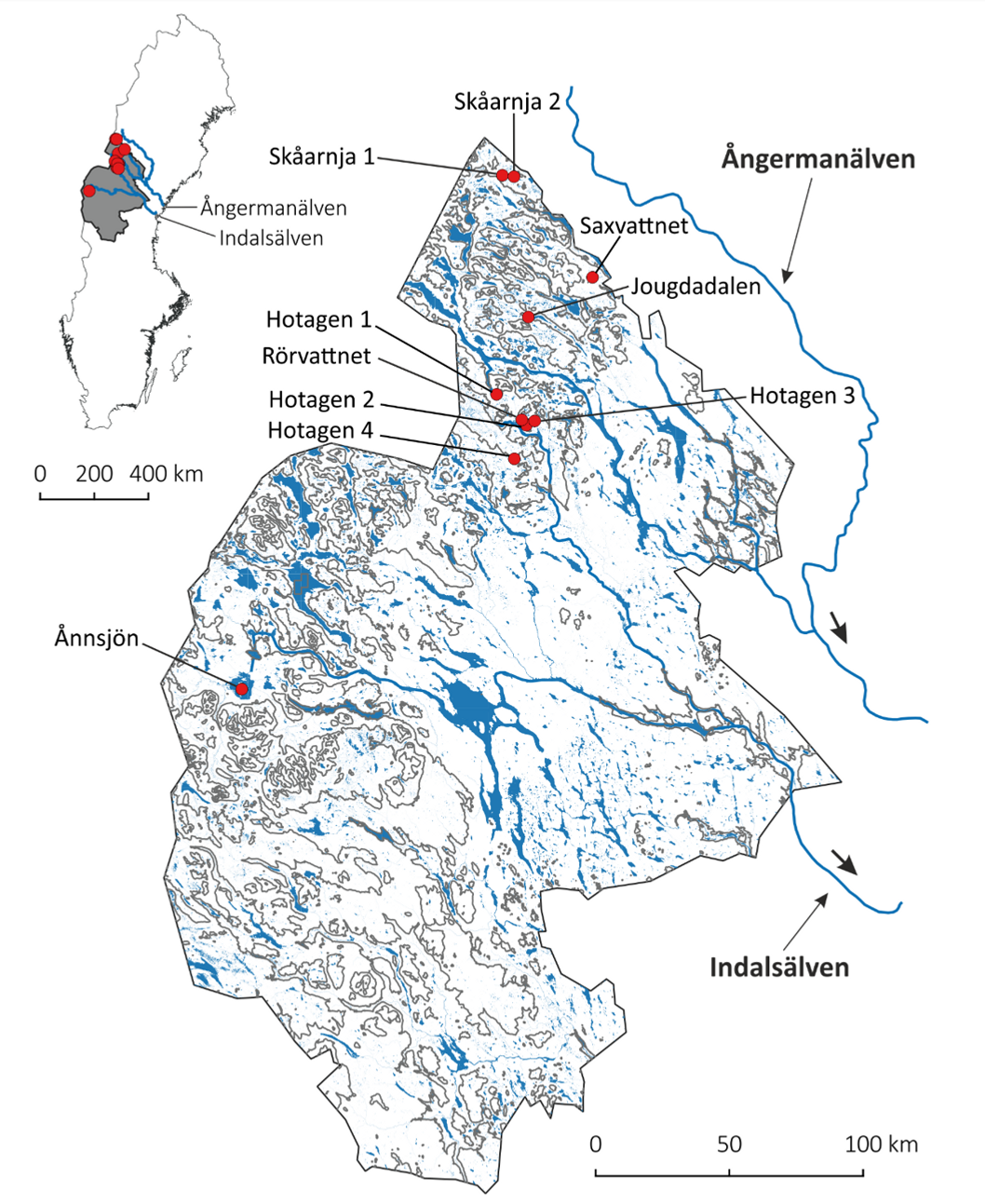
Skyddade områden i Jämtland där genetisk mångfald hos öring har övervakats över perioden 1970-80-tal till sent 2010-tal. Skyddet har införts efter det att första insamlingen gjordes och innan den andra insamlingen gjordes. Övervakningen har skett med SNPs i samtliga områden (en panel med 96 SNP-markörer spridda över arvsmassan). I vissa områden (delar av Skåarnja 1 och 2, Saxvattnet, Hotagen 4 och Ånnsjön har även helgenomsekvensering använts. Resultaten är publicerade vetenskapligt i Andersson et al. (2022) och Kurland et al. (2023).
Övervakningen omfattade att först identifiera antalet genetisk särpräglade populationer inom varje vattendrag/-system/sjö. Därefter undersöktes i vilken utsträckning dessa populationer även kunde identifieras i materialet insamlat ca. 40 år senare i samma vatten. Den genetiska variationen inom och mellan de genetiskt distinkta populationerna mättes med de EBVs som ingår i indikatorerna.
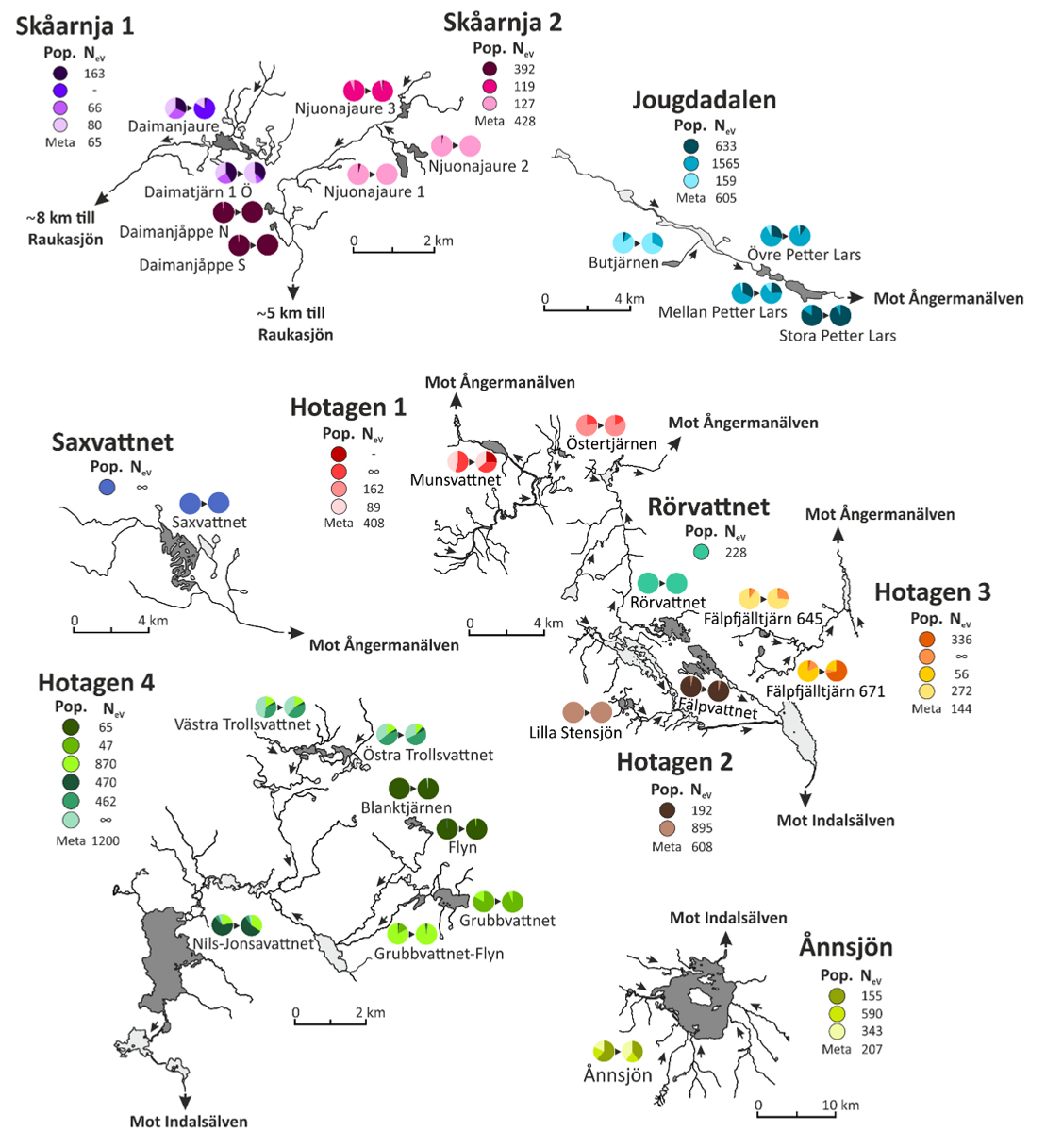
Genetiskt distinkta populationer som identifierats inom de olika vattensystem där den genetiska variationen övervakats från 1970-80-tal till sent 2010-tal (fem-sex generationer). Cirkeldiagrammen anger med hjälp av färger antalet genetiskt distinkta populationer som identifierats i materialet från 1970-80 i diagrammet till vänster samt i materialet från sent 2010-tal. Ingen genetisk population har försvunnit, i tre fall upptäcks en population som inte hittades i det tidiga materialet. Genetiskt effektiv populationsstorlek för varje genetisk population samt för systemet som helhet.
Metapopulationerna Skåarnja 1 och 2 ligger ovanför trädgränsen och i dessa system hittas lägre grad av genetisk variation och till viss del negativa trender över tid.
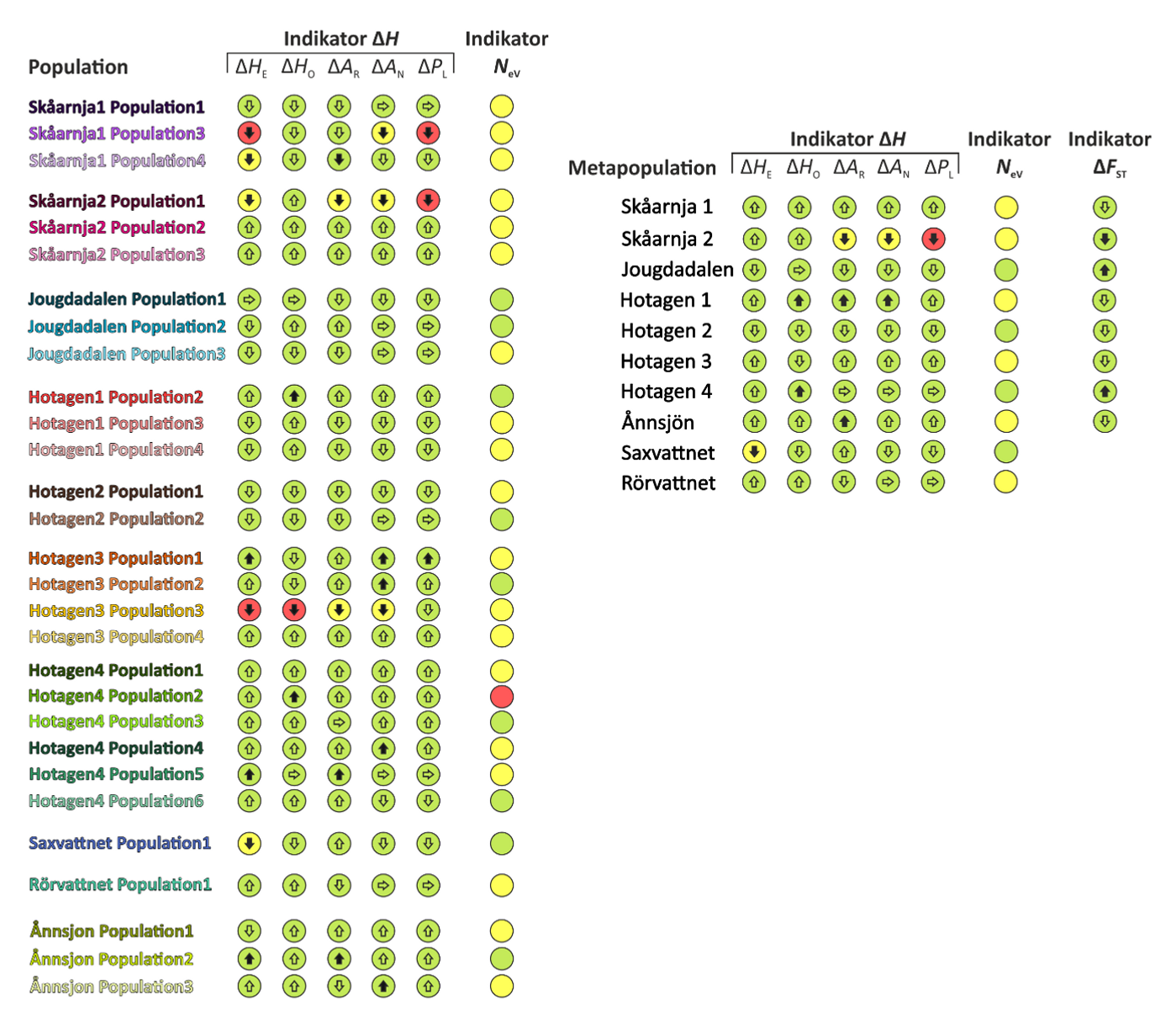
Indikator-utslag avseende genetisk variation för öringpopulationer i fem olika skyddade områden i Jämtlandsfjällen med avseende på indikatorerna ΔH, Ne och ΔFST, baserat på jämförelser mellan två temporala stickprov. De färgade cirklarna visar genetisk status; grön=god/acceptabel, gul=varning och röd=alarm. Pilarna anger den detekterade förändringens riktning, där horisontell pil innebär att ingen förändring har skett; svarta pilar visar på att förändringen är statistiskt säkerställd (p<0.05). Till vänster visas värden för varje genetiskt särpräglad population, till höger för metapopulationerna som helhet. Saxvattnet och Rörvattnet är två sjöar där endast en population per sjö upptäckts.
Varningar och även alarm visas för flera genetiska populationer - totalt 20 av 29 populationer (69%) visar gul varning för Ne indikatorn Ne<500 och i ett fall ligger Ne under 50. Fem populationer av 29 visar även en eller flera varningar för förändringar i EBVs som ingår i ΔH-indikatorn (17%) och i fyra av dessa fall sammanfaller varningar i ΔH med varing för Ne-indikatorn. Tre av dessa populationer ligger ovanför trädgränsen (Skåarnja 1 och 2).
När metapopulationerna beaktas (populationer som hänger samman via genetiskt utbyte) blir läget bättre men 60% har fortfarande Ne<500. Dessa områden är dock inte isolerade utan kontakt finns med ytterligare områden. Vi ser även att ΔH-indikatorn ger utslag för två områden, och i ett fall sammanfaller detta med varningar för låg Ne parallellt med ett statistiskt signifikant minskat genetiskt utbyte (ΔFST) som dock inte är så stort och med nuvarande tröskelvärden klassas minskningen som grön=acceptabel. Denna metapopulation finns ovanför trädgränsen i naturreservatet Skåarnja (metapopulation Skåarnja 2).
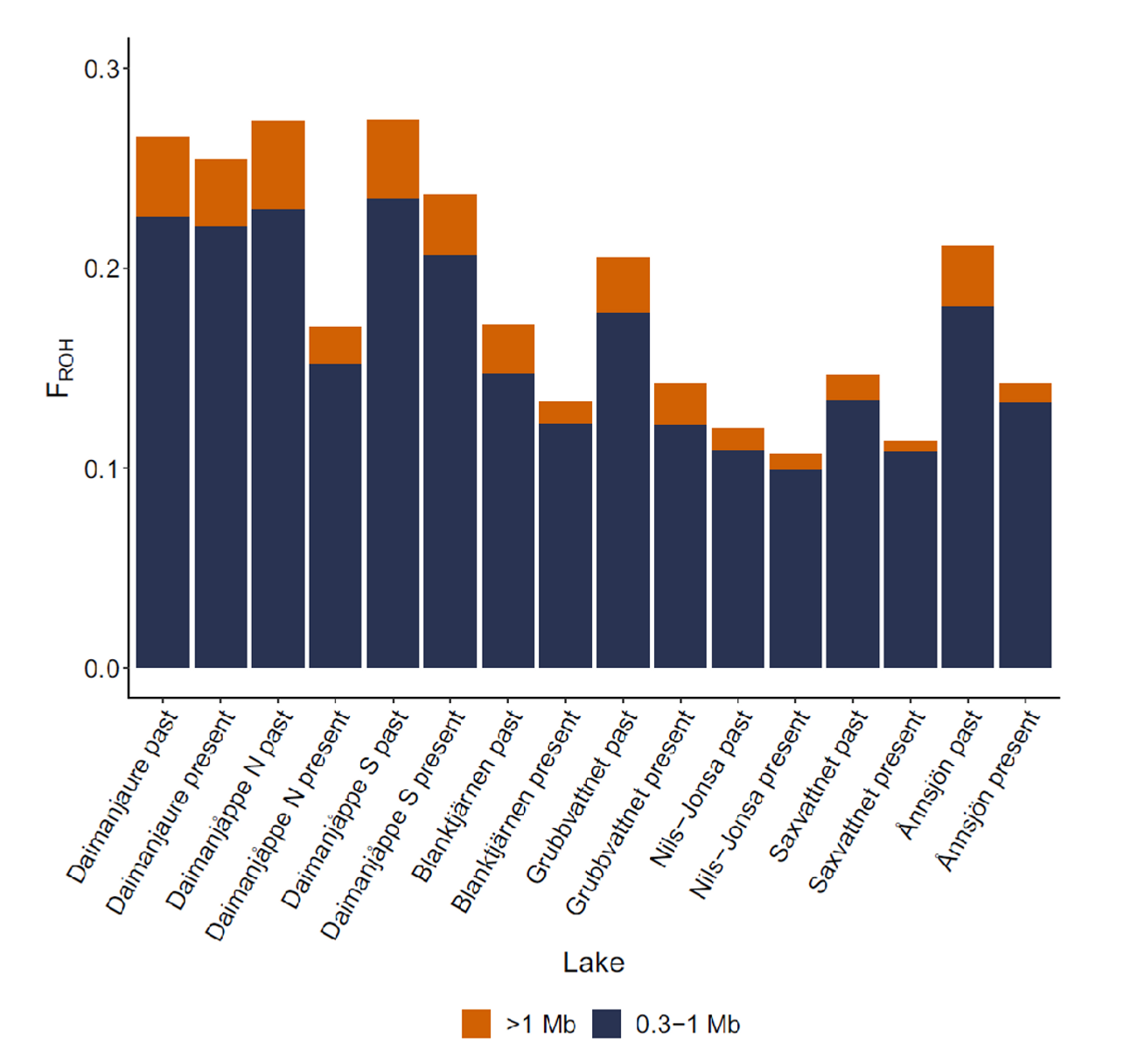
Genomikstudier visar att inavelsnivåerna är högre i populationerna ovanför trädgränsen (de sex första staplarna). Orange visar på mängden ny inavel som genererats över senaste generationerna medan blått indikerar gammal inavel som kan ha byggts upp under lång tidsperiod.
Genomikstudierna identifierar även tecken på selektion över de fem-sex generationerna som sker parallellt över de undersökta sjöarna. Gener som kunna identifieras bland de selekterade ingår sådana som styr pigmentering. I ett vattendrag (Grubbvattnet i Hotagens naturreservat) upptäcks ett så kallat selektivt svep i en kromosom och där gener som är inblandade i reproduktion och immunologi ingår.
Nya analyser
För närvarande pågår analys av havsöring på Gotland samt en mer detaljerad analys av Skåarnja-populationerna.
Frågor av särskild relevans
Öringens tendens att bilda genetiskt särpräglade bestånd även över begränsade geografiska områden bör tas i beaktande i förvaltningen. Vi ser att de genetiskt effektiva storlekarna för enskilda populationer kan vara mycket låga och klart under gränsvärdet Ne>500 i majoriteten av fall. Av de 29 populationer som studerats har 20 st (69%) Ne<500 och en population har Ne<50. När vi däremot studerar sammanhängande metapopulationer (system av populationer där genetiskt utbyte kan ske mellan populationer minskar andelen Ne<500 till 60% och ingen metapopulation visar rött för Ne-indikatorn. Låga värden för flera metapopulationer i kombination med gröna värden för ΔH-indikatorn tyder dock på att genetiskt utbyte sker över större geografiska områden än vad som kunnat övervakas genetiskt. Sammantaget ser vi att möjlighet till genetiskt utbyte över stora geografiska områden är centrala för öring i fjällsjösystem. När sådant utbyte äger rum, som i dessa skyddade områden, nås en god genetisk situation. Sårbarheten är dock tydlig, särskilt i områden ovanför trädgränsen. Där observerar vi en metapopulation som ger varning på både ΔH- och Ne- indikatorn trots att det rör sig om ett skyddat område.
Vill du läsa mer om arten?
Läs mer på sidan Öring.
Vetenskapliga nyckelreferenser
- Andersson A, Karlsson S, Ryman N, Laikre L. 2022. Monitoring genetic diversity with new indicators applied to an alpine freshwater top predator.
Molecular Ecology 31:6422-6439 DOI: 10.1111/mec.16710 - Kurland S, Saha A, Keehnen N, de la Paz Celorio-Mancera M, Díez-del-Molino D, Ryman N, Laikre L. 2023. New indicators for monitoring genetic diversity applied to alpine brown trout populations using whole genome sequence data.
Molecular Ecology 2024;33:e17213. - Saha S, Andersson A, Kurland S, Keehnen NLP, Kutschera VE, Hössjer O, Ekman D, Karlsson S, Kardos M, Ståhl G, Allendorf FW, Ryman N, Laikre L 2022. Whole-genome resequencing confirms reproductive isolation between sympatric demes of brown trout (Salmo trutta) detected with allozymes.
Molecular Ecology 31:498-511. DOI: 10.1111/mec.16252

I Bottenhavet bildar blåstången sammanhängande tångskogar ner till cirka 10 meters djup som är viktiga för smådjur, småfisk och för strömmingens lek. Foto: Ricardo Pereyra.
Tidigare genetisk kartläggning
Den första genetisk kartläggningen (2005) gjordes med en handfull mikrosatelliter och redan detta avslöjade att blåstång kan bilda kloner, vilket inte var känt tidigare. Stora kloner har sedan hittats utmed svenska och finska kusterna av Bottenviken och i Finska viken. Idag visar kraftfulla genetiska analyser att kloning även förekommer på vissa lokaler i Skagerrak-Kattegat men inte alls i den omfattningen som i Bottenhavet och Finska viken. Slutsatsen är att kloning är möjlig överallt men har gynnats i vissa sammanhang. Äggens problem med låg salthalt och en klons fördel vid kolonisering av ett nytt område, är möjliga förklaringar.
Den genetiska variationen hos blåstången i hela dessa utbredning i Östersjön är idag analyserad med kraftfulla genetiska metoder, DNA sekvensering som genererat 37 000 informativa loci (SNPs). Med dessa resultat framstår blåstång som en art med kraftiga genetiska skillnader mellan olika sexuella bestånd i sin utredning från Nordsjön till Bottenhavet. Förekomsten av en handfull stora kloner varav en (smaltång) har miljontals individer utmärker Bottenhavet och Finska viken. Men dessutom finns stora genetiska skillnaderna mellan sexuella bestånd på avstånd ner till kilometer. Dessa reflekterar både anpassning till Östersjöns lokala förhållande, men också barriärer för genflöde och resultat av koloniseringshistorien. De senaste resultaten pekar på att blåstången i Östersjön precis som blåmusslan och Östersjömusslan utgör en egen evolutionärgren som avknoppades från Atlantens blåstång redan långt innan Östersjön bildades.
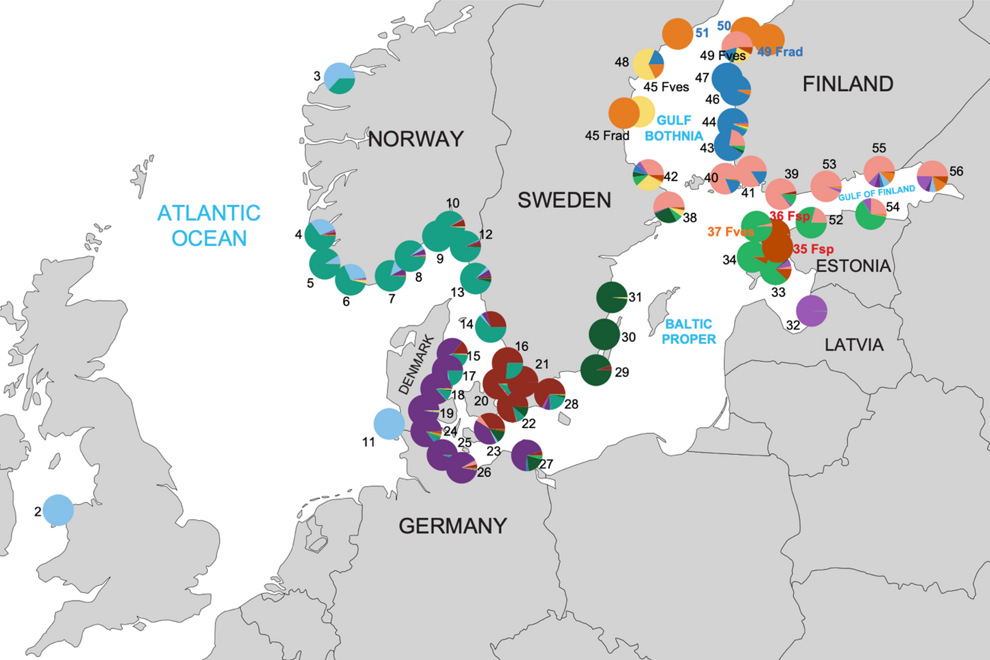
Blåstången är i Östersjön, Öresund, Bälten, Kattegatt och Skagerack uppdelad i genetisk särpräglade bestånd. Detta beror troligen på en kombination av anpassning till brackvatten och historiska förklaringar. Alla västliga lokaler t.o.m. 31 (Västervik) representerar en Atlantisk gren av blåstång. Resterande lokaler inne i Östersjön består av smaltångsklonen (orange färg, lokalerna 42, 45,48-51), den estländska evolutionära grenen (brun färg på lokalerna 35, 36), och resterande Östersjö-gren av blåstång. Illustration ur: Pereyra et al 2025.
Resultat från nuvarande övervakningsprogram
Det saknas historiska vävnadsprover av blåstång men inom ramen för övervakningsprogrammet för genetisk mångfald har prover samlats in från flera lokaler runt hela svenska kusten med ett tidsintervall av åtta år (2014-2022). Dessa prover kan ge en indikation på den temporala dynamiken hos arten. Speciellt intressant är det att följa om klonerna ökar eller minskar i utbredning. Kloner har generellt svårt att anpassa sig genom naturligt urval till pågående miljöförändringar dock kan viss selektion ske mellan asexuell avkomma med olika mutationer.
Frågor av särskild relevans
- Var Östersjöns blåstång har sitt ursprung är i sig intressant men viktigast är att utröna om det sker hybridisering med västerhavets bestånd eller om Östersjöbestånden är isolerade.
- Den lokala enklaven av blåstång i Estland som verkar vara isolerad från övrig blåstång behöver ett starkt skydd för att inte försvinna. Om detta är en för Östersjön endemisk art eller ej behöver utredas. För detta behövs provtagning av bestånd i andra områden med låg salthalt (till exempel inre delar av norska fjordar).
- Inom de områden där den stora smaltångsklonen dominerar finns på vissa platser sexuella populationer. Dessa kan ha väldigt stor betydelse för blåstångens möjligheter att överleva i Bottenhavet och dessa bestånd bör kartläggas och skyddas.
- Då blåstången viktiga funktioner i framförallt Östersjöns ekosystem kan restuarering av förlorade blåstångsbestånd bli aktuella. Metoder för detta behöver tas fram och utvärderas med avseende på genetiska faktorer. Att införa sexuellt förökande individer i områden dominerade av en klon skulle kunna bli aktuellt för att öka potentialen för anpassning.
Vill du läsa mer om arten?
Läs mer om Blåstång hos Artfakta.
Vetenskapliga nyckelreferenser
- Hatchett, W. J., Coyer, J. A., Sjøtun, K., Jueterbock, A., & Hoarau, G. (2022). A review of reproduction in the seaweed genus Fucus (Ochrophyta, Fucales): Background for renewed consideration as a model organism. Frontiers in Marine Sciences 9, 1051838.
En översiktsartikel som beskriver reproduktionsbiologin inom släktet Fucus, och förekomsten av hybridisering mellan många av arterna. - Johansson, D., Pereyra, R. T., Rafajlović, M., & Johannesson, K. (2017). Reciprocal transplants support a plasticity-first scenario during colonisation of a large hyposaline basin by a marine macro alga. BMC Ecology, 17, 14.
En studie som visar att blåstången i Bottenviken är anpassad till extremt låg salthalt och växer bättre i 4 promille än i 24 promille. - Jonsson, P. R., Kotta, J., Andersson, H. C., Herkul, K., Virtanen, E., Nyström Sandman, A., & Johannesson, K. (2018). High climate velocity and population fragmentation may constrain climate-driven range shift of the key habitat former Fucus vesiculosus in the Baltic Sea. Diversity and Distribution, 24, 892–905.
En modellering som utifrån ekologiska parametrar visar hur blåstångens utbredning kommer förändras i Östersjön fram till nästa sekelskifte. - Pearson, G., Kautsky, L., & Serrão, E. (2000). Recent evolution in Baltic Fucus vesiculosus: reduced tolerance to emersion stresses compared to intertidal (North Sea) populations. Marine Ecology Progress Series, 202, 67-79.
En experimentell studie som visar att Östersjöns blåstång är anpassad till att ej torrläggas (vilket västkustbestånden klarar utan problem). - Pereyra, R. T., Rafajlović, M., De Wit, P., Pinder, M., Kinnby, A., Töpel, M., & Johannesson, K. (2023). Clones on the run -- the genomics of a recently expanded partially clonal species. Molecular Ecology, 32, 4209–4223.
En modelleringsstudie som ger en möjlig förklaring till hur de stora klonerna av blåstång har uppstått och spridits i Östersjön. - Pereyra RT, Kinnby A, Le Moan A, Ortega-Martinez O, Jonsson PR, Piarulli S, Pinder MIM, Töpel M, De Wit P, André C, Knutsen H, Johannesson K. 2025. An evolutionary mosaic challenges traditional monitoring of a foundation species in a coastal environment - the Baltic Fucus vesiculosus. Molecular Ecology | Molecular Genetics Journal | Wiley Online Library
En heltäckande studie av den genetiska variation hos blåstång runt svenska kusten. Studien visar bland annat att den tidigare beskrivna arten smaltång (Fucus radicans) är en stor klon av blåstång med en utbredning utefter hela svenska Bottenhavskusten. - Serrão, E. A., Brawley, S. H., Hedman, J., Kautsky, L., & Samuelsson, G. (1999). Reproductive success of Fucus vesiculosus (Phaeophyceae) in the Baltic Sea. Journal of Phycology, 35, 254–269.
Experiment som visar att sexuell förökning försvåras vid låga salthalter pga polyspermi. - Wikström, S. & Kautsky, L. (2006). Structure and diversity of invertebrate communities in the presence and absence of canopy-forming Fucus vesiculosus in the Baltic Sea. Estuarine, Coastal and Shelf Science, 72, 168-176.
Studien visar hur blåstång fungerar som ett viktigt habitat för ryggradslösa djur.

Torsken är bottenlevande men de befruktade äggen är pelagiska. Torsken är en av våra viktigaste rovfiskar. (Torsken på bilden lever i akvarium). Foto: Natalie Greppi / Havs- och vattenmyndigheten.
Tidigare genetisk kartläggning
Torskens tre huvudsakliga bestånd
Atlanttorskens genetiska variation har kartlagts i stora delar av dess utbredning, vilket omfattar hela norra Atlanten. Några av bestånden är migrerande och befinner sig vid olika platser under speciellt lek och uppväxt. Andra bestånd är stationära och ofta lokala. Under senare år har tusentals eller fler genetiska markörer används för att identifiera olika lekbestånd.
Inom den genetiska övervakningen som drivs av HaV används en s.k. SNP panel med cirka 4000 genetiska markörer. Provtagning sker runt hela kusten upp till södra Kvarken. I huvudsak särskiljs tre genetiskt skilda bestånd av torsk. Ett i Östersjön ("östra beståndet"), ett i Öresund, Kattegatt och i Skagerrak fjordar ("kusttorsk" eller "fjordtorsk"), och ett tredje i utsjön i Skagerrak och Kattegatt med huvudsaklig utbredning i Nordsjön ("nordsjötorsk"). Av dessa är det östra beståndet mest genetiskt särpräglat, och utmärks också av en rad anpassningar till Östersjöns lägre salthalt. Forskning visar att det idag inte sker någon hybridisering mellan östra beståndet och den kusttorsk som finns i sydvästra Östersjön (ibland också kallad "västra beståndet"). Biologiskt innebär detta att det östra beståndet är att likställa med en endemisk art av torsk även om en formell artbeskrivning ej gjorts.
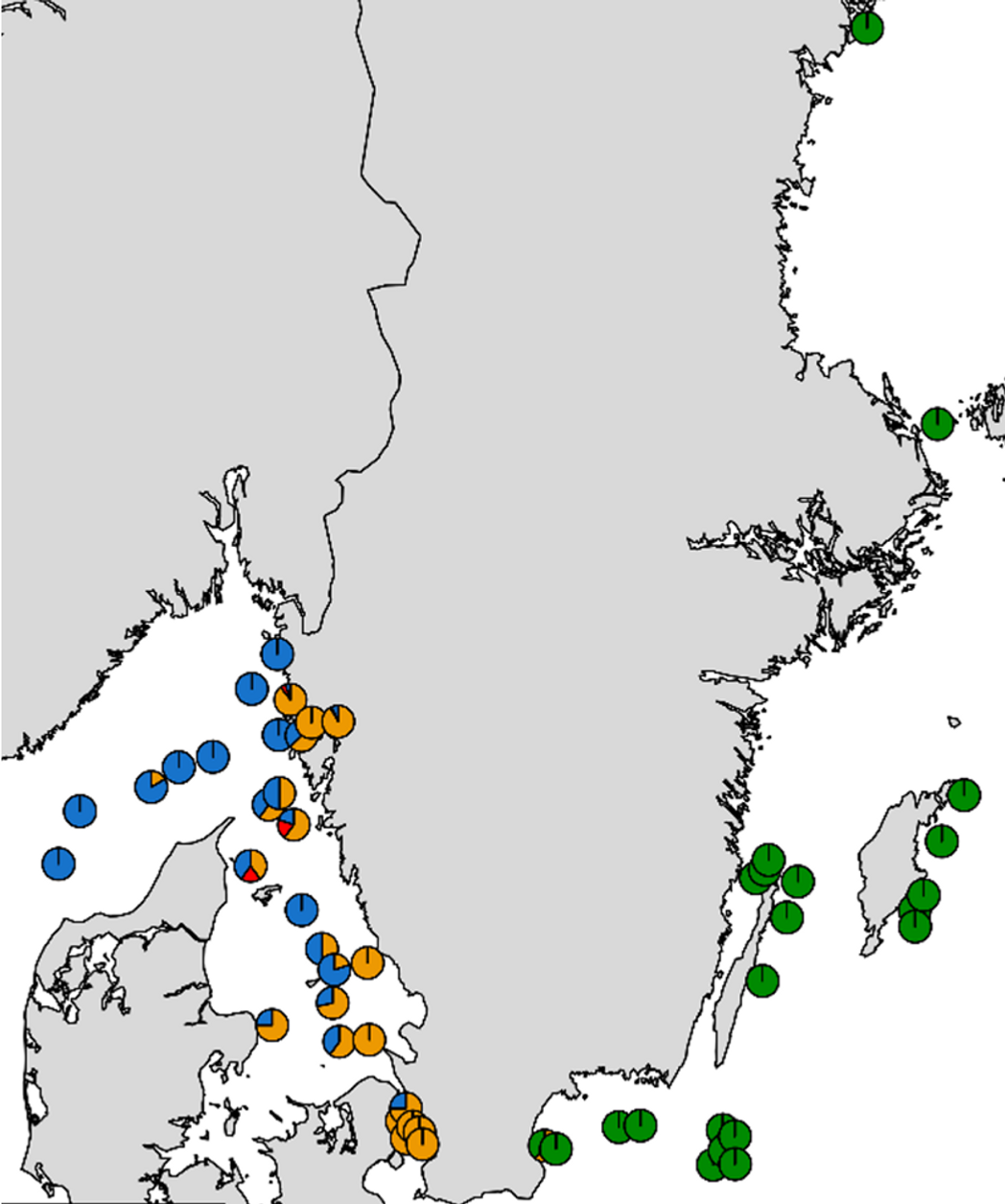
Med ca 4000 genetiska markörer framträder tre olika bestånd, Östersjöns östra bestånd (grön), Öresund/Kattegatt och Bohusläns fjordar (orange) och Nordsjöbeståndet som framförallt leker väster om Danmark men även i utsjön i Skagerrak (blå). Som framfår av kartan överlappar utbredningen av Nordsjöbestånden och Kattegattbeståndet i Kattegatt.
Torsken i västkustens fjordar
Tidigare förekommande torsklek i fjordarna i norra Halland (Kungsbacka fjorden) och i Bohuslän (Hakefjorden, Gullmarsfjorden, Brofjorden, Åbyfjorden m.fl.) har idag i stort sett upphört. Dock återfinns ett fåtal lokalt producerade torskägg i samband med vårleken. Men det finns en stor risk att det rör sig om ett mycket litet antal lekande torskar. Däremot visar genetiska studier av torskyngel att både kusttorsk och nordsjötorsk uppehåller sig i fjordarna under sitt första år. Nordsjöynglen har färdats med strömmarna från lekområdet väster om Danmark, men det är ännu inte fastställt varifrån ynglen av kusttorsk härrör. Antingen är det lokalt producerade eller så har det färdats med strömmarna från södra Kattegatt och Öresund. Studier med så kallad helgenomsekvensering pågår för att få klarhet i dessa yngels ursprung.
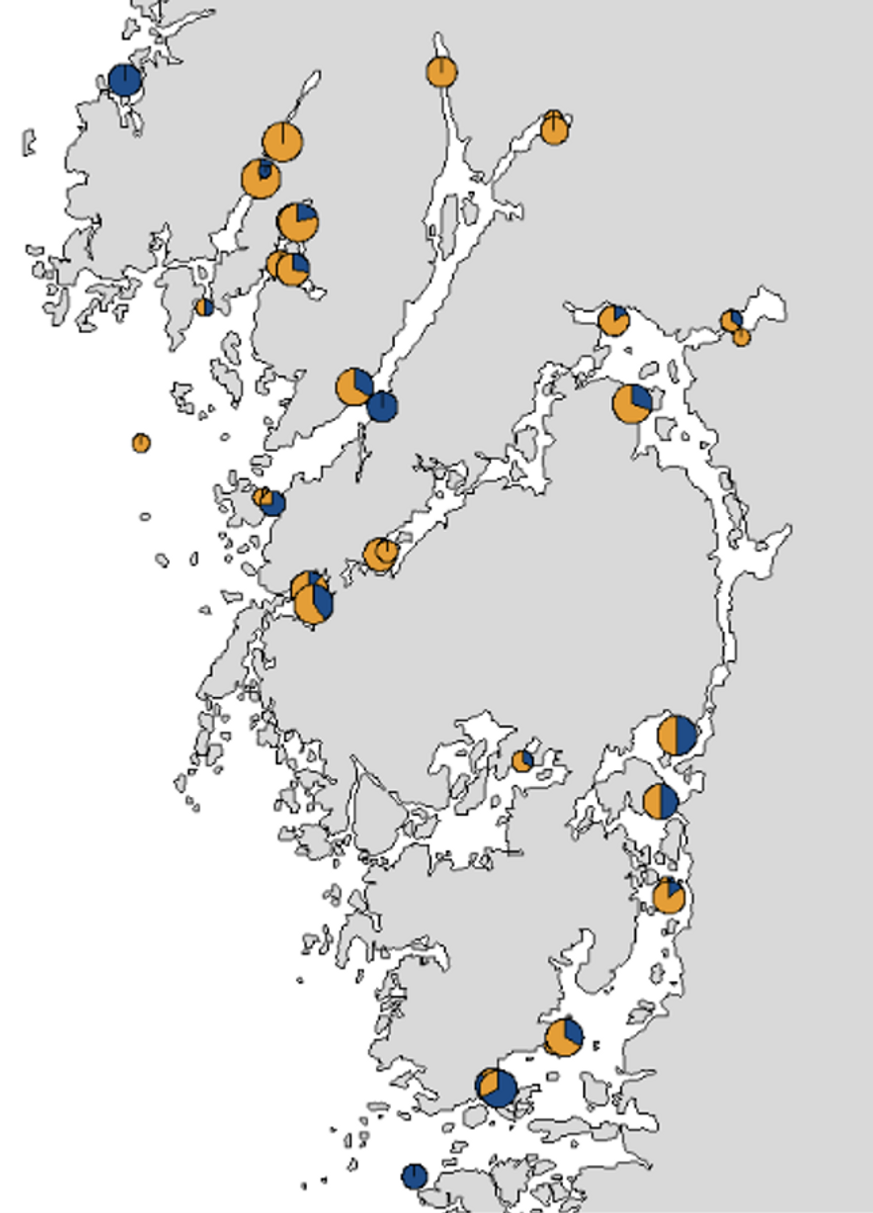
Yngel av kusttorsk (orange) och nordsjötorsk (blå) återfinns båda i i fjordarna runt Tjörn och Orust, och i Gullmarsfjorden, Brofjorden och Åbyfjorden.
Planer inom nuvarande övervakningsprogram
En SNP panel med flera tusen genetiska markörer som skiljer ut alla olika kända bestånd är ett effektivt sätt att genomföra övervakningen. Insamlingen av prover kan samordnas med de regelbundna fiskeribiologiska insamlingarna som sker i svenska vatten inom ramen för annan övervakning.
Frågor av särskild relevans
Det saknas historiska vävnadsprover av torsk och det finns därför ännu ingen data som kan avslöja trender i den genetiska variation över tiden.
Det östra beståndet i Östersjön har en lägre genetisk variation (31.6%) än de två andra bestånden (34.6% och 35.0%). Detta kan vara en effekt av tidigare perioder av populationsflaskhalsar, som till exempel i samband med Östersjöns tillkomst för 8000 år sedan. Det östra beståndets isolering innebär att genetisk variation inte längre tillförs beståndet genom migration av individer från andra bestånd. Det är därför mycket viktigt att försöka behålla så mycket genetisk variation som möjligt i det östra beståndet genom att behålla ett så stort bestånd av reproducerande individer som möjligt. Övervakning behövs som syftar till att bevaka vad som händer med den genetiska variation över tid. Idag (2023) har Östersjön östra bestånd ännu en effektiv populationsstorlek (Ne=3700) som är tillfredsställande ur perspektivet att beståndet ännu inte riskerar genetisk utarmning. Dock är en så stor genetisk variation som möjligt alltid en fördel för att stärka ett bestånds potential att klara miljöförändringar.
Om det förekommer lek i Ålands skärgård och i Bottenhavet, och om eventuella lokalt lekande individer redan har etablerat ett eller flera lokala bestånd behöver dessa bestånd följas upp med regelbunden genetisk övervakning.
De tidigare stora och livskraftiga bestånden av kusttorsk i Öresund och i södra Kattegatt är idag starkt decimerade. Det finns härmed en ökad risk att genetisk variation går förlorad. Det är mycket viktigt att med genetisk övervakning följa utvecklingen över tid i dessa bestånd och genomföra relevanta åtgärder för att säkra att så stor genetisk variation som möjligt bibehålls för att öka beståndens potential att motstå miljöförändringar.
Om genetiska analyser visar att det fortfarande förekommer lokal lek av kusttorsk i någon/några av fjordarna på västkusten (undersökning pågår 2024-2025), behöver dessa lekområden skyddas och bestånden skyddas så kraftfullt att de får en positiv utveckling. Genetisk övervakning bör användas för att följa upp skydds och restaureringsåtgärder.
Det förekommer olika åtgärder för restaurering av torskbestånd både på västkusten och i Östersjön. Artificiella rev har etablerats på flera platser i Bohuslän. Både på ostkusten och västkusten förekommer mer eller mindre omfattnde utsättning av torsk. Alla typer av restaureringsåtgärder bör följas upp med noggranna genetiska utvärderingar som utvisar om, och hur, åtgärderna förändrar genetisk variation och genetisk struktur. Det är viktigt att framhålla att omfattande stödutsättningar kan ha negativa konsekvenser för de vilda bestånd man önskar stödja.
Vill du läsa mer om arten?
Läs mer på sidan Torsk.
Vetenskapliga nyckelreferenser
- Barth et al. 2019. Disentangling structural genomic and behavioural barriers in a sea of connectiity. Mol. Ecol. 28:1394-1411.
Denna genetiska studie visar hur starkt genetiskt särsräglat det östra Östersjöbeståndet av torsk är, samt att vissa fjordar i norska Skagerrak har en blandning av tre olika genetiska bestånd, nordsjötorsk, kusttorsk och ett lokalt fjordbestånd. - Helmerson et al. 2023. Evidence of hybridization between genetically distinct Baltic cod stocks during peak population abundance(s). Mol Ecol. 16:1359-1376.
Studien visar att när bestånden av torsk i söda Östersjön var betydligt störr än idag skedde också hybridisering mellan kusttorsken i västa Östersjön-Öresund och torsk från det östra beståndet i Östersjön. Idag finns inga tecken på att ett genutbyte sker med det östra beståndet, vilket poängterar problemen med hög isolering av detta bestånd. - Henriksson et al. 2023. Mixed origin of juvenile Atlantic cod (Gadus morhua) along the Swedish west coast. ICES J. Mar. Sci. 80:145-157.
Genetiska analyser visar att noll-åriga juveniler av torsk som befinner sig i fjordarna i söda Bohuslän består av en mix av kusttorsk (antingen lokalt eller från södra Kattegatt/Öresund) och Nordsjötorsk. Längre in i fjordarna och i de grundare vattnen dominerar kusttorskjuveniler, medan Nordsjötorskjuveniler företrädesvis befinner sig längre ut och på djupare vatten. Men visst överlapp förekommer. - Matschiner et al. 2022. Supergene origin and maintenance in Atlantic cod. Nat. Ecol. Evol. 1:469-481.
Genetisk data från båda sidor Atlanten visar att det finns så kallade supergener (kromosomala inversioner) som kodar för migrationsbeteenden och anpassningar till antingen kustnära liv för torsken eller ett mer migrerande och oceaniskt livsstil. Supergener skiljer kusttorsk och nordsjötorsk från varandra. Även det östra beståndet i Östersjön skiljs ut från övriga bestånd i en av dessa supergener.
Lagring och åtkomst av data från övervakningen
Genetisk inomartsvariation övervakas genom insamling av vävnad från ett representativt antal individer per art, varefter den genetiska sammansättningen fastställs för varje individ. De primära mätdata som genereras används för att beräkna indikatorer som är viktiga för det akvatiska förvaltningsarbetet, och resultaten rapporteras för respektive art. I avsaknad av nationellt datavärdskap kommer både mätdata och de beräknade indikatorerna att lagras hos Havs- och vattenmyndigheten, HaV, och göras fritt tillgängliga för nedladdning. För närvarande pågår utvecklingen av formatet för dessa primära mätdata. När formatet har fastställts kommer information om hur och var dessa data kan laddas ner att publiceras.
Vävnadsprovbanker
Den vävnad som samlas in från individer av de arter som övervakas lagras i vävnadsprovbank där detta är möjligt. Proverna kan lagras i djupfryst tillstånd eller i etanol. Vissa typer av vävnad kan även lagras i torkat tillstånd (främst fiskfjäll och hörselstenar, otoliter). Dessa samlingar är centrala att säkerställa eftersom utvecklingen av nya DNA-metoder går väldigt fort. Att kunna extrahera DNA från äldre vävnad för analys med nya metoder är ständigt aktuellt. Mycket av den övervakning som HaV bedriver är baserad på samlingar från tidigare forsknings- och övervakningsinsatser.


