Uppföljning
%20fiskpassager-passageeffektivitet.jpg)
Foto: Håkan Carlstrand
Den grundläggande funktionen i fiskpassagen är beroende av att konstruktionen är hel, att anlockningsströmmen når ut till fiskarna i miljön nedströms och att tillräckligt med vatten strömmar genom fiskpassagen för att upprätthålla det vattendjup och den strömhastighet som behövs för målarterna. Vid inventeringar av fiskpassager i norra och södra Sverige bedömdes bland annat funktionen. Låg funktion härleddes ofta till bristande underhåll vilket många gånger lett till att vattendjupet var för lågt (Broman 2018, Nilsson 2019). Verksamhetsutövaren ansvarar för att kontrollera och underhålla passagen så att funktionen upprätthålls, till exempel genom att se till att rätt mängd vatten tappas vid rätt tidsperioder, att flödet regleras vid behov, att passagen och eventuella galler rensas från skräp, att felaktigheter åtgärdas etc. Skötsel och tillsyn för att upprätthålla funktionen bör beskrivas i en enkel manual för att underlätta för den ansvarige verksamhetsutövaren och öka chansen till funktionella passager.
Efter anläggandet behöver åtgärden utvärderas för att ge förutsättningar för förbättringsåtgärder (Roscoe and Hinch 2010). Nyvunnen kunskap genom utvärdering och förbättringsarbete bör spridas för att driva den generella utvecklingen framåt. I detta kapitel beskrivs vilka metoder som kan vara relevanta för uppföljning av passageåtgärden, när de är tillämpbara, vilken osäkerhet som kan förväntas, samt hänvisning till litteratur där metoderna beskrivs mera utförligt. Kapitlet berör inte verksamhetsutövarens egenkontroll, men en del av metoderna som beskrivs skulle även kunna användas för det ändamålet.
Förhoppningen är att den tilltänkta passageanordningen leder till att de delar av målbilden som har att göra med konnektivitet uppfylls. Enda sättet att få klarhet i om detta blir fallet är att följa de förändringar i miljön som väntas med lämpliga metoder och med tillräcklig dimensionering (replikering) av datainsamlingen i tid och rum. Med otillräcklig dimensionering försvåras möjligheten att statistiskt påvisa en förändring även om den finns där.
Det kan bli fyra olika resultat av en uppföljning:
- Den förväntade förändringen kan påvisas statistiskt och den är sann.
- Den förväntade förändringen påvisas statistiskt, men förändringen är inte sann. Det fel som begås kallas typ-I fel och beror på slumpen och förekommer med samma sannolikhet som den valda signifikansnivån (a), till exempel a=0,05.
- Ingen förändring kan påvisas statistiskt trots att det finns en verklig förändring. Det fel som begås kallas typ-II fel och beror på otillräcklig datainsamling i tid och rum i relation till variationen i data och beroende på effektens storlek. Risken att begå ett typ-II fel betecknas med symbolen b. Vid dimensionering av provtagningsprogram används begreppet statistisk styrka (1-b).
- Ingen förändring kan påvisas vilket leder till en korrekt slutsats när det inte finns någon effekt att påvisa. Det finns två tänkbara förklaringar till frånvaron av effekt. Antingen beror det på att passageanordningen inte fungerar som den ska, eller så har den förväntade effekten felbedömts. För att skilja mellan dessa två alternativ kan passageanordningens funktion kontrolleras med oberoende metodik.
En grundförutsättning för att kunna göra en korrekt tolkning av en uppföljning är att den förväntade effekten är välgrundad, vilket per definition innebär att omständigheterna är kända innan målbilden formulerades. Om det mot förmodan saknas information om de variabler som ska följas upp behöver data samlas in innan åtgärden genomförs för att användas som referensdata i de kommande utvärderingarna.
Det finns två enkla fall där den förväntade effekten är given. Det ena fallet gäller uppföljning av effektiviteten i olika delar av passagelösningen och det andra fallet gäller uppföljning som handlar om att påvisa kolonisation av de arter som saknas uppströms, eller i sällsynta fall nedströms. Beskrivningen av de uppföljningsmetoder som är aktuella för dessa två fall ges tillsammans med övriga uppföljningsmetoder längre fram i detta kapitel. I vissa fall kan det räcka med kvalitativa beskrivningar av de förväntade effekterna av passagelösningen, men utan kvantifiering kan det hända att t ex en förväntad ökning av ett fiskbestånd blir så pass liten att den inte går att påvisa trots att uppföljningen görs enligt regelboken. För att vara säker på att den förväntade effekten är påvisbar behövs någon typ av kvantifiering av förväntade effekter. Av den anledningen ges en kortfattad beskrivning i nästa avsnitt hur kvantifiering av förväntade effekter kan göras. Om den förväntade effekten av konnektivitetsåtgärden är stor finns också goda möjligheter att kunna påvisa förändringen statistiskt med hjälp av den eller de uppföljningsmetoder som är mest lämpad för ändamålet. Om den förväntade effekten är liten kan det behövas omfattande provtagning för att påvisa förändringen.
Kvantifiering av förväntade populationseffekter för att kunna följa upp effekter av åtgärden
I samband med uppföljning mäts en eller flera variabler i syfte att få kvantitativa mått för att uttala sig om passageåtgärdens effekt på miljön. Utan kunskap om vilka kvantitativa effekter en passageåtgärd kan tänkas få går det inte att veta vilket resultat som krävs i samband med en uppföljning för att visa att åtgärden har avsedd effekt. Ett undantag är dock målarter som saknas uppströms. I dessa fall ger en kvalitativ bekräftelse på etablering ett entydigt svar. Cooke and Hinch (2013) påpekade behovet av kunskap hur till exempel reproduktion eller överlevnad påverkas av passage och vad det får för konsekvenser för populationen. Populationsmodeller som utvecklats för att skatta populationsstorlek och göra prognoser för olika arter kan modifieras för att kvantifiera de förväntade populationseffekterna innan man fastställer vad som ska följas upp.
Vandring, spridning och genetisk diversitet och struktur påverkas av konnektivitet och populationseffekterna kan kvantifieras med populationsekologiska respektive -genetiska modeller. Det är därför möjligt att på förhand få en uppfattning om hur stort bidrag var och en av dessa aspekter av konnektivitet skulle kunna tänkas ha för de enskilda arterna/bestånden som finns i de anslutande vattenförekomsterna, med respektive utan konnektivitet. I samband med modelleringen går det även att få en uppfattning om vilken tidsperiod som krävs innan förändringarna når en storleksordning som gör dem möjliga att påvisas statistiskt (se t ex FiMod 1 & 2, energiforsk.se).
En grundförutsättning för att individerna som vandrar eller sprider sig till nya miljöer genom att passera fiskpassagerna ska kunna göra bättre ifrån sig jämfört med om de hade stannat, är att det finns bättre livsmiljöer att komma till där de kan fullfölja sin livscykel. Det finns i dagsläget endast ett fåtal beräkningsmodeller som utvecklats för att analysera dessa specifika aspekter. För fiskarter där täthetsberoendet (populationsregleringen) sker innan den första vandringen finns tre olika beräkningsverktyg (FiMod1–3). Dessa modeller har hittills främst använts för att utvärdera de förväntade ekologiska konsekvenserna av fiskpassager och deras effektivitet för lax och öring. För lax och vandringsöring yttrar sig täthetsberoendet främst som dödlighet innan nedströmsvandringen. Vandringen från yngeluppväxtområdet till födosöksområdet behöver inte vara speciellt lång, som hos lax, för att modellen ska vara användbar. Modellen fungerar även för de fall när fiskarna, t ex öringar, har korta säsongsmässiga födosöksvandringar. Givet att antagandet om täthetsberoendet gäller, är dessa modeller applicerbara även för andra arter. En anledning till att modellerna tillämpats för lax och öring är framförallt att kunskapen är större om dessa arter vilket gör det lättare att parameterisera modellerna (ange värden för bärkapacitet, överlevnad, fekunditet, passageeffektivitet, etc.) på ett tillförlitligt sätt.
I dagsläget saknas beräkningsverktyg för att utvärdera de förväntade effekterna av fiskpassagers effektivitet på populationsstorleken för arter med pelagiska larver som driftar/simmar nedströms redan inom några dagar/veckor efter utkläckning och passerar hindren innan täthetsberoendet inträffar. För dessa bestånd kan täthetsberoendet antingen yttra sig i form av dödlighet eller reducerad tillväxt/fekunditet. Exempel på arter som hör till denna kategori är harr, sik, mört, stäm, löja, abborre, asp.
Kvantifiering av spridningens betydelse för fiskbestånden går att göra för att utvärdera de populationsgenetiska effekterna med hjälp av individbaserade modeller. Däremot är det betydligt svårare att kvantifiera effekterna av spridningens betydelse för återkolonisation av områden där det lokala beståndet slagits ut. Sannolikheten till utslagning, vanligen till följd av slumpmässig demografisk eller miljömässig variation, är som störst i små bestånd. Huvudorsaken till att dessa bestånd är små torde vara brist på lämpliga livsmiljöer. I detta sammanhang innebär det brist på strömsträckor eftersom sjöliknande miljöer i form av sel och dammar ökat på bekostnad av strömmiljöerna som en följd av vattenkraftsutbyggnaden. De arter som därför har störst återkoloniseringsbehov är strömlevande arter som öring, lax, harr och elritsa.
Det pågår forskningsprojekt för att utveckla fler beräkningsverktyg i stil med FiMod som är lämpade för kvantifiering av såväl populationsekologiska som -genetiska effekter för fler arter och för flera olika typer av vandring än de befintliga modellerna hanterar.
Val av uppföljningsmetoder
Grundtanken med uppföljning i den yttre miljön är att kunna visa att åtgärden har avsedd effekt. En komplicerande aspekt i sammanhanget är om miljön inte återhämtar sig som förväntat. Beror det då på brister i anläggningen, att åtgärdens förväntade effekter felbedömts, eller att man får fel bild av effekterna genom ett otillräckligt uppföljningsprogram? Det är viktigt att inse att alla uppföljningsmetoder inte har möjlighet att svara på samtliga förändringar i miljön som beror på förändrad konnektivitet. I detta avsnitt ligger därför fokus på vägledning för att identifiera en eller flera lämpliga uppföljningsmetoder samt på analys av vad som krävs för att kunna påvisa förändringar med dessa metoder. I Figur 1 sammanfattas information om olika metoder. Genom att följa nedanstående flödesschema (Figur 1) får man en uppfattning om vilka uppföljningsmetoder som kan vara tillämpliga för respektive målart beroende på vilka omständigheter som råder. Det finns dock fler möjliga uppföljningsmetoder än de som listat i flödesschemat, men merparten av de som inte namnges i listan är metoder som kräver speciella förhållanden, som exempelvis räkning av fisk i samband med snorkling. I de fall det finns kompletteringsmetoder som kan öka precisionen i skattningen nämns dessa i metodavsnitten nedan. I Figur 1 ges en översikt av tillgängliga metoder för uppföljning av bland annat fiskpassager. För mer utförlig beskrivning av metoder för uppföljning av åtgärder i vattendrag se Nilsson (2013), och Karlsson och Leonardsson (2014) .
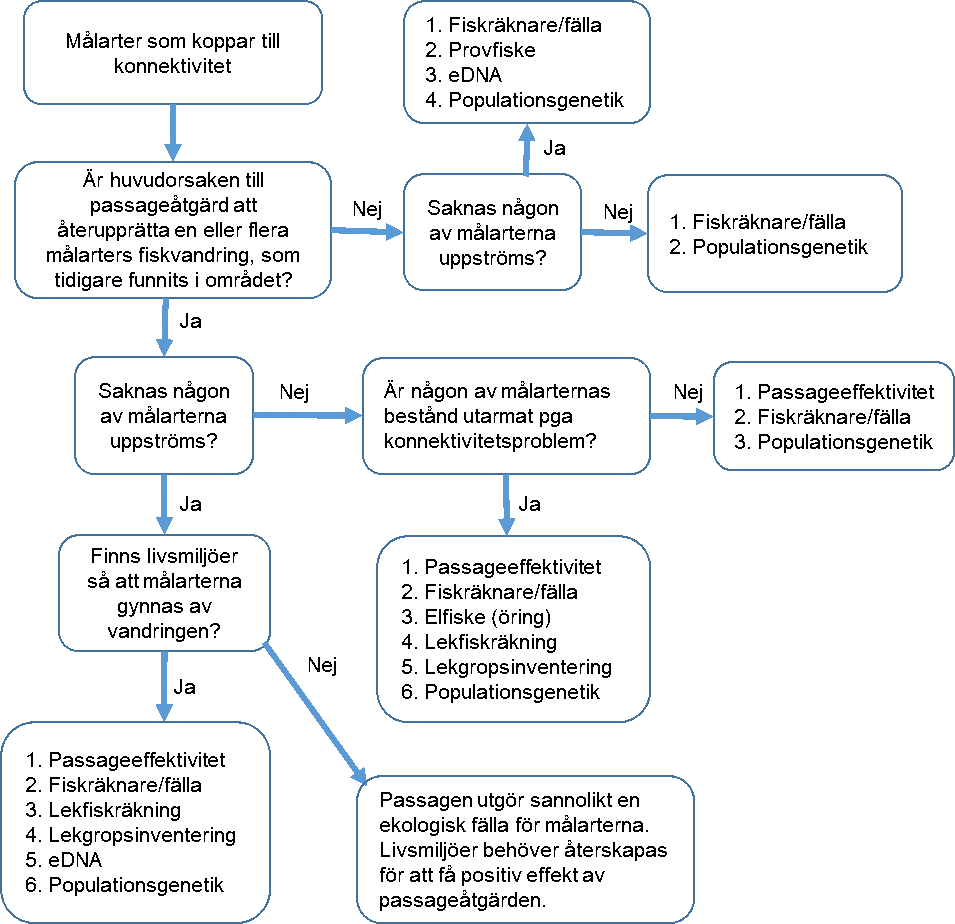 Förstora bilden
Förstora bildenFigur 1. Genom att följa flödesschemat för varje enskild målart får man fram förslag på metoder som kan användas vid uppföljning.
För att underlätta utvärdering och möjliggöra jämförelser är det viktigt att standardiserade metoder används. Detta har även föreslagits i vetenskaplig litteratur där man sammanställt kunskapsläget och framtidsutsikter inom forskningsfältet (Silva med flera 2018), eller försökt göra mer detaljerade meta-analyser av passageeffektivitet i olika fiskpassagestyper och för olika arter (Bunt med flera 2012).
Uppföljningens utformning kan variera från fall till fall beroende på omständigheterna. Uppföljningen kan handla om ett eller flera av följande alternativ:
- undersöka passageeffektivitet förbi anläggningen för målarter med utpräglat vandringsbeteende i området,
- undersöka anläggningens förmåga att anlocka fisk till passagen i uppströms riktning och att avleda fisk i nedströms riktning,
- räkna och dokumentera hur många individer av respektive art som passerar anläggningen på daglig basis i både uppströms och nedströms riktning,
- undersöka förekomst av målarter uppströms med hjälp av eDNA
- undersöka genetisk diversitet/struktur hos målarterna uppströms respektive nedströms för att validera att den genetiska diversiteten ökar (eller strukturen förändras) till följd av passageåtgärden
- för att följa upp förändring i status enligt vattenförvaltningen”, så överensstämmer det med kraven på övervakning i HVMFS 2015:26 avseende relevanta kvalitetsfaktorer
- dokumentera förekomst av lekande vandringsöring eller andra arter genom lekgropsinventering eller lekfiskräkning
- följa upp rekryteringsframgång av t ex vandringsöring genom elfiske. En annan provfiskemetod för att undersöka återetablering av utslagna arter skulle kunna vara nätprovfisken. En nackdel med den metoden är att den är destruktiv och på så vis motverkar sitt eget syfte i detta sammanhang. Dessutom är metoden selektiv och lämpar sig främst för arter som rör sig mycket. Av dessa anledningar rekommenderas inte nätprovfiske vid uppföljning av konnektivitetsåtgärder.
Tabell över användningsområden och tidsperspektiv på uppföljningen kopplat till val av metod för uppföljning
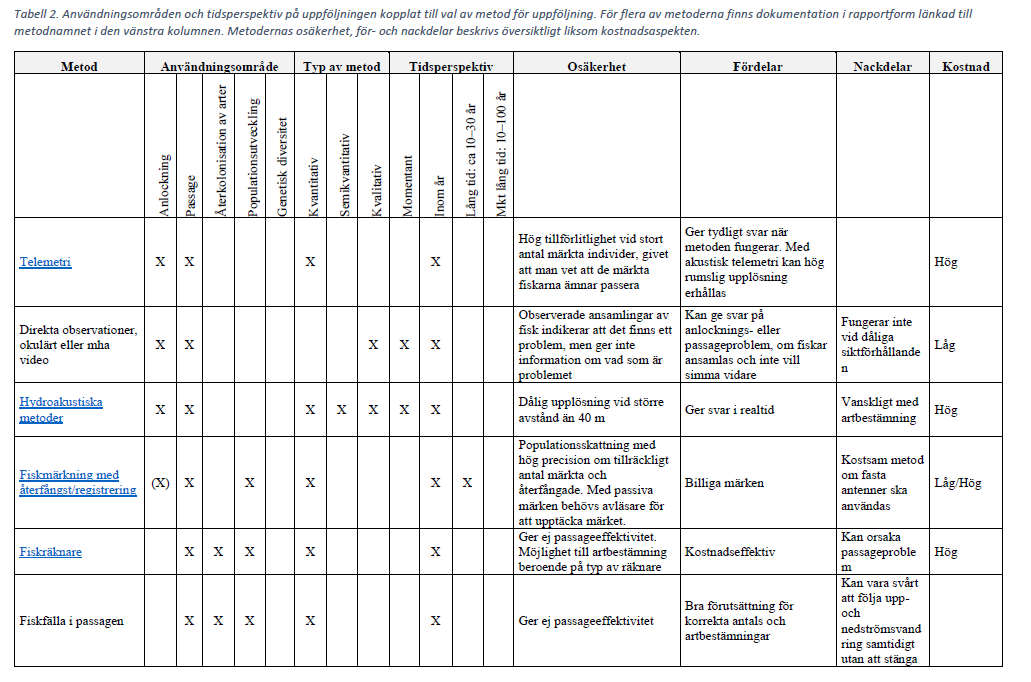
Klicka på tabellen för att visa i större format.
Passageeffektivitet
Det pågår ett arbete med att ta fram gemensamma standarder för att utvärdera fiskpassagelösningar med hjälp av telemetri och dessa förväntas bli färdiga inom kort. Arbetet i Sverige drivs av Svenska Institutet för Standarder (SIS) och bygger på den europeiska standarden: ”Water quality – Guidance for assessing the efficiency and related metrics of fish passage solutions using telemetry” (prEN 17233). Här anges standardiserade metoder för att utvärdera fiskpassagelösningar med hjälp av telemetri både för uppströms- och nedströmslösningar. I den europeiska standarden ingår även rekommendationer av utrustning, studiedesign, dataanalys och rapportering.
Passageeffektivitet som uppföljningsmetod är begränsad till ett fåtal arter och situationer och utvärderingen ger endast svar som rör passageanläggningens funktion. Den effektivitet som behövs i en enskild fiskpassage för att uppnå målbilden beror av de biologiska förutsättningarna, var fiskpassagen är placerad i avrinningsområdet och hur många fler passager som vandringsfiskarna behöver passera innan de når målhabitatet. Generellt gäller att ju längre nedströms i vattensystemet desto viktigare med hög effektivitet, liksom vid flera på varandra följande passager (Porcher and Travade 2002). Då en fiskpassage i en del fall endast anlockar fisk från en del av vattendragets bredd kommer den, även med optimal funktion, att fungera som en flaskhals. Detta betyder att ingen fiskpassage kan förväntas erbjuda konnektivitetsaspekter i samma utsträckning som innan vandringshindret uppfördes (DWA 2014). Om huvudsyftet med passagen är att möjliggöra genetiskt utbyte mellan populationer uppströms och nedströms, räcker det dock med att ett fåtal individer passerar per generation som är framgångsrika i reproduktionen (Hanski and Gaggiotti 2004).
Undersökningar för att skatta passageeffektivitet (se faktaruta) syftar till att ge tillförlitliga svar som kopplar till olika delar av anläggningen, när man vet vilka individer som ämnar passera anläggningen men som misslyckas på grund av anläggningens utformning eller funktion. I fiskpopulationer där till exempel alla lekfiskar vandrar från område (A) till ett annat område (B) finns goda förutsättningar att utvärdera passageeffektiviteten i en anläggning som finns däremellan. Det blir inte lika enkelt att tolka resultaten om endast en del av populationen ämnar ta sig till område B eftersom det kräver att man känner till individernas avsikt (Kemp 2016). I vissa situationer skulle det kunna vara möjligt, men det kräver i så fall att individer som ska till område B skiljer sig genetiskt från de som inte ska dit. En utvärdering av passageeffektivitet baserat på individer från den senare typen av populationer blir därför mycket svårare och kostsammare att utföra och utvärdera.
En kategori fiskar som är direkt olämpliga att använda för utvärdering av passageeffektivitet är de som saknar ett utpräglat vandringsmönster. Individer tillhörande denna kategori kan mycket väl passera genom fiskpassager, men ofta mer slumpmässigt fördelat över säsongen. Utvärdering av passageeffektivitet baserat på individer av den sistnämnda kategorin kan i dessa fall endast genomföras med ett fåtal individer som dessutom utgör en liten del av beståndet. Risken är dock stor att den verkliga passageeffektiviteten underskattas i sådana situationer.
Under förutsättning att det saknas lek- och yngeluppväxtområden för öring nedströms passagen skulle det vara möjligt att utvärdera passageeffektiviteten för uppströmsvandrande leköring. Om det å andra sidan är möjligt för öring att leka nedströms passagen kan en passageeffektivitetsstudie med märkt fisk leda till underskattning av passageeffektiviteten eftersom en del av de märkta individerna kanske inte hade för avsikt att passera även om de utforskade hela området upp till fiskpassagen innan de bestämde sig. Inom forskningen om passageeffektivitet har man på senare tid börjat dokumentera och mäta motivationen hos fiskarna (Goerig and Castro-Santos, 2017), vilket kan ge ökade möjligheter till uppföljning av passageeffektivitet för andra arter än öring och lax. För nedströmsvandrande öring (kelt och smolt) och ål går det att utvärdera passageeffektiviteten eftersom dessa behöver passera för att komma till havet. Däremot torde en utvärdering av passageeffektivitet för arter som abborre, mört, sutare och gädda ge en direkt felaktig bild av möjligheten för dessa arter att anlockas och passera anläggningen eftersom man inte vet om de märkta individerna avser att passera. Även om dessa bestånd tidigare har haft uttalad vandring i det aktuella området är det inte nödvändigtvis alla individer i populationen som vandrat.
Även om kriterierna för utvärdering av passageeffektivitet är uppfyllda kan det uppstå situationer när passageeffektiviteten underskattas, t ex om konditionen eller hälsan inte är fullgod hos de individer som märks, alternativt att hanteringen i samband med märkningen påverkar motivationen att fortsätta vandringen. I de vetenskapliga publikationerna som rör passageeffektivitet ägnas litet utrymme i diskussionen åt att hitta andra förklaringar till låga passageeffektivitetsvärden än att hänvisa till bristande funktion i anläggningen.
Anlockning/avledning
Utvärdering av anlockningsförhållandet kan omfatta kartläggning av anlockningsplymens storlek och avstånd från konkurrerande flöden från anslutande damm eller kraftverk, till exempel med hjälp av drönare. Om djupet i anlockningsområdet är tillräckligt och vattenmassan är fri från luftbubblor kan det gå att använda sonar/ekolodsteknik för att få kunskap om i hur stor utsträckning fiskarna i området kommer i kontakt med respektive attraheras av anlockningsströmmen. Luftbubblor, även små sådana om de förekommer i stor mängd, hindrar ljudsignalerna att nå ut i vattenmassan. Metoden kan ge svar på hur många fiskar som uppehåller sig i området, hur stora de är och hur de rör sig i förhållande till anlockningsplymen. Tekniken fungerar ungefär som en videokamera men bilden återskapas av reflektioner av ljudsignaler som sänds ut och studsar tillbaks till kameran, som ett ekolod. Skillnaden mot ett vanligt ekolod är att resultatet som erhålls med en mer avancerad sonar är mer likt en videosekvens där man kan se fiskarna i svartvitt. Det går även att få information om avstånd till fiskarna och deras storlek och den är inte beroende av ljusförhållandet. Det går att särskilja en del arter om de är stora, men små fiskar torde vara svåra att artbestämma. En viss återkoppling mellan fiskarna man ser i sonarutrustningen borde gå att få genom att jämföra med fiskar som registrerats i fiskpassagen med hjälp av fiskräknare eller fiskfälla i passagen.
Metoden ger svar med hög upplösning om fiskarna visar intresse för anlockningsvattnet och hur de beter sig vid avledningsanordningar. Metoden har samma begränsningar som telemetristudier genom att det inte går att veta vilka fiskar som är motiverade att passera anläggningen. En nackdel med metoden är att utrustningen är dyr, ca 1 mkr, och att genomgång och analys av ”filmerna” är tidskrävande om inspelningarna täcker flera veckor. Analyskostnaden kan dock reduceras om man fokuserar på stickprov snarare än på hela det inspelade materialet.
Räkning av passerande fisk
Oavsett om det finns målarter med utpräglat vandringsbeteende i området eller inte kommer troligen merparten av arterna som passerar anläggningen att förekomma i lågt antal. Passagen för den senare kategorin ger dock möjlighet till genetiskt utbyte och eventuell återetablering av utslagna bestånd. Med automatiska fiskräknare och fiskfällor får man dokumentation om antal individer av respektive art som passerar i uppströms respektive nedströms riktning. Om det finns flera passageanläggningar i nära anslutning efter varandra i ett vattendrag skulle det kunna räcka med fiskräkning i den första och den sista anläggningen. Det skulle göra det möjligt att få ett grovt mått på passageeffektiviteten. Om det visar sig att passageeffektiviteten är låg kan man flytta räkningen av fisk från den övre passagen till den närmast nedströms för att på så vis ringa in passageproblemet.
För att kunna utvärdera antal uppströms- respektive nedströmspassager av olika arter som registrerats i automatiska fiskräknaren behöver man utesluta registreringar av individer som tvekar kring räknaren, exempelvis simmar fram och tillbaks en eller flera gånger innan den bestämmer sig. Den information som behövs är om individen som passerar kom från vattenförekomsten uppströms i samband med nedströmsvandring eller om individen kom från vattenförekomsten nedströms i samband med uppströmsvandring. Detta torde endast vara möjligt att urskilja fiskar med jojo-beteende om det är få fiskar i rörelse och tidsskillnaderna mellan uppströms- och nedströmsregistreringarna är små. Detta problem uppstår inte när man använder fiskfällor för att räkna antal passerande fiskar. Slutredovisningen ska innehålla separata kolumner för uppströms- respektive nedströmspassager för att kunna avgöra hur många individer som passerat i endera riktningen.
Det är lämpligt att resultaten skickas till en datavärd efter avslutad säsong, efter att kvalitetsgranskning gjorts av data. Det finns en ”preliminär” metod för att bedöma passageeffektivitet för fiskräknardata där det framgår vad man ska tänka på i samband med kvalitetssäkring av såväl teknik som analys och kvalitetssäkring av data från fiskräknare (framtagen av KAU på uppdrag av HaV). SLU kommer att slutföra metoden för att bedöma passageeffektivitet under 2020 och kommer dessutom att vara datavärd för data från fiskräknare.
eDNA
Med hjälp av DNA-analyser av filtrerade vattenprov går det att fastställa vilka arter som finns i vattendraget. Det går att få en viss uppfattning om den relativa mängden av de olika arterna genom att många individer släpper ifrån sig mer DNA än få individer. Kvantifieringen är dock osäker, men metoden kan ge svar på om en art förekommer inte alltför långt uppströms provtagningsplatsen. Vid väldigt låg förekomst av en art ökar risken att artens DNA inte kommer med i analysen. Därför kan det behövas prov från några olika platser i vattenförekomsten. Metoden är relativt kostsam, men tids- och kostnadseffektiv jämfört med metoder som baseras på insamling av fisk. Metoden är fortfarande under utveckling vilket innebär att en del av de osäkerheter som finns med metoden sannolikt kommer att avhjälpas inom de närmaste åren. Naturvårdsverket finansierar för närvarande flera forskningsprojekt med inriktning mot eDNA och fisk.
Med hjälp av DNA-analyser av filtrerade vattenprov går det att fastställa vilka arter som finns i vattendraget. Det går att få en viss uppfattning om den relativa mängden av de olika arterna genom att många individer släpper ifrån sig mer DNA än få individer. Kvantifieringen är dock osäker, men metoden kan ge svar på om en art förekommer inte alltför långt uppströms provtagningsplatsen. Vid väldigt låg förekomst av en art ökar risken att artens DNA inte kommer med i analysen. Därför kan det behövas prov från några olika platser i vattenförekomsten. Metoden är relativt kostsam, men tids- och kostnadseffektiv jämfört med metoder som baseras på insamling av fisk. Metoden är fortfarande under utveckling vilket innebär att en del av de osäkerheter som finns med metoden sannolikt kommer att avhjälpas inom de närmaste åren.
Naturvårdsverket finansierar för närvarande flera forskningsprojekt med inriktning mot eDNA och fisk.
Populationsgenetik
Fragmentering av miljön som innebär frånvaro av konnektivitet gör att fisken av olika anledningar kommer att påverkas. Genetiska förändringar uppstår som ett resultat av genetisk drift, mutationer, naturligt urval (selektion) och genflöde. Genetisk drift är en slumpmässig process som beror på att ett begränsat antal individer för sina gener vidare vid varje generationsskifte. I små populationer går genetisk drift snabbare än i stora populationer. Vid inrättande av dammar minskar strömhabitatet och populationsstorlekar för de fiskarter som är beroende av dessa. Således ökar risken för genetisk driften och även att vissa genetiska egenskaper försvinner ur små populationer. Dessutom kan selektionstrycket förändras, exempelvis för vandringsbenägenhet. Den mest uppenbara konsekvensen av bristande konnektivitet är minskat genetiskt utbyte (genflöde) mellan populationer, eller inom populationer i samma vattendrag (vilket leder till fragmentering). Genetiskt utbyte kan vara viktigt för långsiktig livskraftiga populationer då detta kan motverka negativa effekter av den genetiska driften.
Med hjälp av genetiska analyser går det att studera förändringar som uppstår till följd av mänskliga aktiviteter, exempelvis fragmentering eller minskade habitatytor. De centrala och vanligast förekommande uppföljningsmåtten är genetisk diversitet (eller variation), genetisk struktur, och effektiv populationsstorlek. Till detta kan nämnas att man med hjälp av genetiska analyser kan identifiera enskilda individer, till exempel strayers, och beräkna graden av migration (genflöde) över tid. I det sammanhanget kan det vara bra att känna till att populationsgenetiska studier skiljer sig från kvantitativa populationsstudier, av antal individer, genom att resultaten analyseras och tolkas mot bakgrund av existerande genetiska teorier och hypoteser medan antalsskattningar inte påverkas av ekologiska teorier. Populationsgenetiska studier har stor potential att ge värdefull information men den snabba kunskapsutvecklingen inom ämnet gör det svårt för andra än aktiva forskare att utforma relevanta provtagningsprogram, analysera och tolka resultaten.
Det mått på genetisk diversitet som i nuläget rekommenderas i litteraturen för analys av fragmenterade populationer är alleldiversitet (variation av anlagsvarianter) (López-Cortegano med flera 2019), som vanligtvis uttrycks som förväntad heterozygotigrad (He) eller allelic richness (AR). Förändringen av genetisk diversitet i stora populationer går långsamt, men kan gå snabbare i väldigt små populationer (till exempel kraftigt påverkade), särskilt om det öppnas vandringsvägar till populationer som varit isolerade under lång tid. Då kan förändringar i diversitet upptäckas direkt efter lyckad reproduktion, under förutsättning att beståndet som varit isolerat hunnit utsättas för genetisk drift. Generellt sett kan det krävas 10–20 generationer innan förändringar av diversitet får genomslagskraft. De flesta sötvattensarter i Sverige har en generationstid på 2–6 år, vilket innebär att uppföljningen kan behöva fortgå i 20–100 år för att vara säker på att påvisa förändring i genetisk diversitet. En eventuell uppföljning av fiskpassageeffekter avseende genetisk diversitet är därför inte nödvändig med årlig provtagning, (även om det kan krävas för stabila skattningar av effektiv populationsstorlek), utan vart femte eller vart tionde år kan vara mer lämpligt. Diversitet (He) kan ge en bild av hur känslig en population är för ytterligare förändringar i miljön. För en art som öring kan ett diversitetsmått ge en bild av hur stort utbyte som sker till exempel med havsöring. Generellt sett har sötvattenlevande öring lägre diversitet (He ~0,57) än havsvandrande öring (He ~0,66) (Söderberg med flera 2017, Östergren and Nilsson 2012).
Genetisk struktur analyseras genom att studera relationen mellan två eller flera populationer, eller genetiska stickprov. Här används måttenheten FST (FST definieras som den proportion av den totala genetiska variationen hos två eller flera stickprov som förklaras av skillnader mellan stickproven; (Weir and Cockerham 1984)) för en generell (global) bild av strukturen, men också för parvisa jämförelser. Man kan till exempel jämföra skillnader i FST mellan uppströms och nedströms (vandringshinder) och studera parvisa förändringar över tid. Genetisk struktur kan illustreras genom så kallade dendrogram (släktträd), eller PCA (principal component analysis). Det finns även dataprogram som analyserar ett eller flera genetiska prov och delar upp dessa i en eller flera genetiskt homogena grupper baserat på populationsgenetiska teorier (exempelvis Hardy-Weinbergs lag), (t.ex. Structure; (Pritchard and Wen 2004). Struktur och parvisa skillnader kan med fördel användas för att relatera just de ”egna” proven med data för andra områden, vilket ger en bättre bild av vad man kan förvänta sig i effekt av en inrättad fiskpassage (Dannewitz med flera 2012, Dannewitz med flera 2014, Dellefors and Dannewits 2007).
Ett annat mått som ofta används i populationsgenetiska sammanhang kopplat till bevarandeaspekter är effektiv populationsstorlek. Tidigare angavs att den genetiskt effektiva populationsstorleken per generation (Ne) bör vara minst 50 för att undvika inavelsrelaterade problem i ett kortare perspektiv (till exempel (Allendorf and Ryman 2002)). Senare rön tyder dock på att Ne > 100 kan utgöra en mer lämplig riktlinje för att reducera inavelsrelaterade problem i en isolerad population över kortare tidsperioder (Frankham med flera 2014). Ne kan även användas tillsammans med He för att beräkna förväntad förlust av genetisk variation över tid, vilket har använts för att studera effekter av kraftverksdammar (till exempel (Dellefors and Dannewits 2007)).
Det finns olika sätt att beräkna effektiv populationsstorlek och det pågår fortfarande forskning inom området (Hössjer med, flera 2016, Olsson and Hössjer 2015, Waples med flera 2014). Fiskar har överlappande generationer och en del arter har dessutom en komplex livscykel vilket komplicerar beräkningsproceduren och även tolkning av resultatet.
Det finns ett antal aspekter att tänka på viktiga för populationsgenetiska uppföljningsstudier, förutom det långa tidsperspektivet som nämns ovan. För att kunna göra analyserna måste fisk samlas in, helst på ett icke destruktivt sätt. Insamlingen måste även vara representativ, dvs. hela populationen måste vara adekvat provtagen. Olika typer av analyser kräver dessutom att man tar hänsyn till eventuell familjestruktur (Waples and Anderson 2017, Östergren med flera 2019). Det är mycket viktigt med information om sådant som kan ha påverkat den genetiska sammansättningen som inte har med fiskpassagen att göra, till exempel fiskutsättningar, försurning och fiske. Genetiska effekter kan vara svåra att koppla till funktion och behov av förbättringsåtgärder i en nyanlagd fiskpassage med tanke på det förhållandevis fåtal individer som krävs för att upprätthålla den genetiska diversiteten i kombination med det långa tidsperspektivet, även om det går att identifiera (och kvantifiera) antal genetiskt ”nya” individer som passerar en fiskväg. Genetiska studier är enda sättet att bekräfta att ”nya” individer reproducerar sig framgångsrikt i nya tillgängliga habitat. Eftersom resultaten från populationsgenetiska undersökningar är beroende av hur proverna samlas in rumsligt och tidsmässigt är det inte självklart att utformning av provtagningen överensstämmer med andra provtagningsmetoder. För att öka chansen att få svar på de frågor som ställs är det bättre att utforma provtagningen specifikt för ändamålet än att kompromissa och riskera att resultaten inte blir tillförlitliga.
Provfiske
Uppföljning och utvärdering av den ekologiska statusen/potentialen utförs i nuläget vart sjätte år inom ramen för vattenförvaltningen för att kunna verifiera om och när den uppsatta miljökvalitetsnormen nås. Metoderna för denna uppföljning beskrivs i Havs- och vattenmyndighetens bedömningsgrunder och föreskrifter (statusklassning och MKN: HVMFS 2019:25, krav på övervakning HVMFS 2015:26). Nackdelen med denna uppföljningsmetod är att det förväntas ta många år, cirka 10–30 år, innan effekten av fiskpassagen får full genomslagskraft i fisksamhället. En komplicerande faktor vid uppföljning av miljönyttan av en enskild fiskpassage är att miljönyttan inte behöver uppstå i vattenförekomsten där fiskpassagen finns.
De bedömningsgrunder för fisk som skulle kunna vara aktuella är för fisk i sjöar (kapitel sex i HVMFS 2019:25) och för fisk i vattendrag (kapiterl sju HVMFS 2019:25). Provtagningsmetoden för fisk i sjöar baseras på nätprovfiske medan den i vattendrag baseras på elfiske. Bedömningsgrunden för fisk i vattendrag förutsätter att det finns eller har funnits naturlig förekomst av öring och/eller lax i vattendraget samt att det finns lämpliga lokaler för elfiske. Om det saknas övervakningsdata för att klassificera fisk i vattendrag ska i första hand ett underlag samlas in. Om det inte är möjlighet att tillämpa bedömningsgrunden för fisk i vattendrag, beroende på begränsningarna som anges ovan, kan det vara nödvändigt med en expertbedömning. Då kan den hydromorfologiska bedömningsgrunden för konnektivitet i uppströms-nedströms riktning användas som stöd. Den senare baseras på andelen av vandringsbenägna arter (enligt referensförhållandet) som finns uppströms jämfört med nedströms hindret där passagen anläggs. En uppföljning med bedömningsgrunden för konnektivitet kan därför göras genom att utvärdera resultaten från en fiskräknare/fiskfälla och jämföra med referensförhållandena för att fastställa om de saknade arterna passerat uppströms. Ett alternativ till fiskräknare/fiskfälla för just denna typ av uppföljning vore att använda sig av eDNA.
Om det finns underlag för att visa att det är befogat att använda elfiske som uppföljningsmetod (Undersökningstyp: vadningselfiske) uppstår frågan hur många elfiskelokaler som behövs för att kunna påvisa den förväntade effekten. Med få lokaler blir medelvärdesskattningen osäker vilket gör det svårt att kunna påvisa en förändring trots att den finns där. I ett uppföljningsprogram eftersträvas därför hög statistisk styrka. Ett sätt att öka den statistiska styrkan är att öka antalet provtagningslokaler. Av kostnadsskäl görs ofta en avvägning där 80 procent statistisk styrka har blivit något av en norm. För att få en uppfattning om hur många provtagningslokaler som krävs för att kunna påvisa en minsta förändring, givet 80 procent statistisk styrka, har data från elfiskeregistret (SERS) analyserats (Figur 2). Med ett tiotal lokaler krävs att passageåtgärden ger en ökning av öringtätheten (0+) med minst 40–100 procent för att kunna påvisas när data från två olika år jämförs. Om data istället samlas in årligen under en tolvårsperiod kan den första sexårsperioden testas mot den andra sexårsperioden vilket med 5 lokaler per år ger möjlighet att påvisa en förändring som är minst 20–40 procent. För att elfiskemetoden ska vara användbar som uppföljningsmetod för att koppla resultaten till passageanläggningens funktion krävs därför att den förväntade förändringen uppgår till minst 30–65 procent. För öring går det att kvantifiera den förväntade effekten i förväg, vilket också gör det möjligt att dimensionera provtagningen noggrannare än att välja något av utfallen i exemplen ovan. För exempel på hur dimensioneringsberäkningar kan göras se till exempel Leonardsson och Blomqvist (2012), Leonardsson med flera (2016) och Leonardsson och Blomqvist (2018).
Ett sätt att öka precisionen i täthetsskattningen från elfisket vore att märka de fångade fiskarna som sedan återutsätts. Med ytterligare ett fångsttillfälle någon dag senare går det att kombinera utfiskningsmetoden med märkning-återfångstmetoden och på så sätt få en mer tillförlitlig skattning av populationstätheten. Om man använder märkning–återfångstmetoden går det dessutom att få tillförlitliga skattningar för fler arter än öring, om än begränsat till grunda strömvatten.
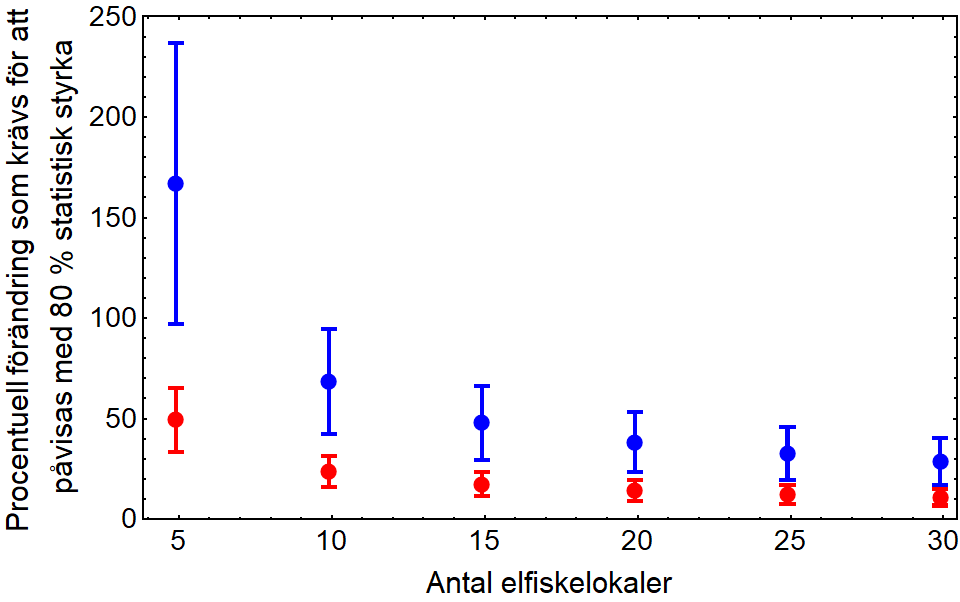 Förstora bilden
Förstora bildenFigur 2. Antal elfiskelokaler som krävs för att påvisa med 80 % statistisk säkerhet att en viss procentuell förändring av täthet av öring (0+) har skett. Blå symboler visar parvisa t-test mellan två olika år och röda symboler visar parvisa test för två 6-års perioder med årliga elfisken. Osäkerhetsintervallerna ligger mellan 25:e och 75:e percentilen.
Lekgrops- och lekfiskinventering
Dessa två metoder baseras på räkning och dokumentation med hjälp av observationer i fält. Metoderna är relativt billiga att utföra. Båda metoderna avser främst Lekgropsinventeringen gäller främst öring, lax och flodnejonöga och kan dokumenteras för att få kvantitativa mått. Lekfiskinventering är mer kvalitativ och kan tillämpas på asp, harr, lax och öring. Se Meddelande nr 2013:18, Länsstyrelsen Jönköpings län


